
3 システム神経科学と精神医学の基本原則
脳は複雑ですが、絶望的ではありません
精神疾患の根底にあるメカニズムを理解する際の大きな困難は、これらの疾患が人体の最も複雑な器官に影響を及ぼし、その器官の最も複雑な側面、つまり私たちを最も人間らしくしている脳の特徴を変化させることです。私たちは、脳がどのように組織され、思考、感情、モチベーションがどのように生成されるのかについて、まだ理解の始まりにすぎません。したがって、精神医学は神経学よりも科学の発展においてはるかに原始的な段階にありますが、2 つの分野の間には明らかに重複する領域があります。実際、神経学には、第 2 章で概説した認知 (認知症) 疾患や、さまざまな形態のてんかんや外傷性脳損傷など、行動神経学の一部と考えられるいくつかの疾患など、少なくとも精神医学と同じくらいよく理解されていない領域があります 。興味深いことに、これらは神経学と精神医学に多くの重複がある分野です。
脳は複雑であると言っても過言ではありません。合理的な推定によると、人間の脳には少なくとも 1,000 億 (10^11) のニューロンがあると考えられています。これらの細胞は入力を処理し、出力を作成し、脳の計算の主力となります。また、各ニューロンには 100,000 (10^5) 以上の接続 (シナプス) がある可能性があります。他のニューロンと結合し、その結果、直接的なニューロン結合の合計数は約 10^15 ~ 10^16 になります。この数は、どの定義から見てもほとんど理解できない驚異的な数ですが、通常、ニューロンをサポートしていると考えられている細胞であるグリア細胞がこれをさらに複雑にしています。グリア細胞、特にアストロサイトは、ニューロンにエネルギーを供給し、ニューロンと協力してニューロン間の伝達を媒介する方法を制御します。また、グリア細胞は、グリア伝達と呼ばれるプロセスを通じて神経伝達物質を放出し、神経活動を調節することができるため、脳の計算作業にも関与します。
この膨大な数の細胞と結合がどのように機能して思考、感情、行動を生み出すのかを理解するのは気が遠くなる作業です。おそらくこの複雑さのため、また精神医学の臨床的側面は臨床神経科学の「よりソフトな」、あまり明確に定義されていない側面を扱っているため、ほとんどの精神科医は脳が精神病理学にどのように寄与しているかを議論する際に間接的で比喩的な概念を使用しているようです。 「よりソフトに」というのは、精神医学的問題が現実ではないということを意味するのではなく、それらが特定の配線された脳経路との直接的な結びつきが少なく、脳の病理があまり明確ではないということだけを意味します。これらの特徴により、通常、神経学は扱いやすくなり、精神医学の観点からはあまり興味のない研究分野になります。
脳の複雑さに対処するために、精神科医の中には(少数派が減っていくことを願っていますが)努力を放棄し、脳を「ブラックボックス」として扱う人もいます。このような概念的なアプローチでは、入力が到着し、「ブラック ボックス」によって何らかの不可解な方法で処理され、その結果、観察可能な出力 (スピーチ、行動、気分など) が得られます。これらの精神科医や他のメンタルヘルス専門家は、機能不全を引き起こす結果とその結果に対して何をすべきかに焦点を当てています。脳内で行われる処理についてはほとんど考慮されていません。このようなアプローチは、自我心理学、抑圧された記憶、行動主義に大まかに基づいた特定のアプローチなど、思考や行動を説明する方法について、善意はあってもよく考えられていない多数のアイデアを生み出す可能性があります。 「ブラック ボックス」アプローチの主な問題は、脳がどのように思考、感情、行動を生み出すかについての基礎が欠如していることです。
多くの「生物学的精神科医」が使用する別のアプローチは、脳を「化学器官」として扱い、脳の機能、病気、治療を化学用語で説明することです。このアプローチの利点は、ニューロンとグリアがメッセージの送受信に神経化学物質を使用するという事実に対処できることです。また、精神薬理学的治療がよく知られた神経伝達物質系の活動を操作し、有益な反応をもたらす可能性があるという事実を利用します。精神科医によく知られている神経伝達物質の例には、人間の脳の主要な高速興奮性伝達物質であるグルタミン酸が含まれます。ガンマ-アミノ酪酸(GABA)、主要な高速抑制性伝達物質。そして生体アミン(ドーパミン、ノルエピネフリン、セロトニン)です。生体アミンはよく知られた名前になっており、脳の領域内および領域全体の活動を調整するのに役立つ重要な調節因子です。それらはまた、多くの向精神薬の主要な標的でもあります。 「化学臓器」のアプローチは、必ずしも正しいとは限りませんが、精神障害の原因についていくつかの興味深い仮説を導き出しました。例としては、「統合失調症のドーパミン理論」、「うつ病のカテコールアミン理論」、「うつ病と不安症のセロトニン理論」などがあります。このアプローチの弱点は、これらの理論が脳が実際にどのように行動、思考、感情を生成するかをあまり効果的に説明していないことです。さらに、このアプローチは、精神疾患が「化学的不均衡」から生じる、または「シナプスの漏洩」の産物であるという、実際よりも深遠に聞こえる概念である、時代遅れの考え方を含め、病気を過度に単純化して説明する傾向があります。
私たちの前提は、精神科医がシステムとネットワーク神経科学に精通し、現代の神経科学が精神科患者に提供する可能性をこの分野で活用するために、神経システムが思考、感情、動機を生成する方法をよりよく知らなければならないということです。このレベルで脳を理解することは、脳システムがどのように機能し、どのように接続されているかを理解する手段として前述した化学器官仮説と一致します。この情報を伝え、概念化する上での問題は、精神科医が知る必要がある詳細レベルです。脳領域間の接続をマッピングした脳配線図は非常に難解であり、「すべてがすべてに接続されている」という絶望的な考えにつながる可能性があります。これはあるレベルでは真実であるように見えますが、ニューロンがどのように機能し、主要な回路がどのように動作するかを扱う中間的な戦略が、知的絶望を克服するのに役立つと私たちは信じています。内因性接続ネットワーク (ICNS) の動作と病態生理学に関する証拠は山積しており、出発点として最適です。
ネットワーク理論と脳システム
ICN について詳しく説明する前に、現代のネットワーク理論と、生物学的システムがどのように組織されていると考えられているかについて少し理解することが重要です。このような背景は、神経システムがどのように機能するかを考えるための概念的な足場を提供します。この議論は、私たちを「スモールワールド」ネットワークと「スケールフリー」ネットワークの科学に導きます。この研究から導き出された原理は、遺伝子やタンパク質から神経結合や高次の神経系に至るまで、さまざまな生物学的ネットワークに適用できるようです。おそらくさらに驚くべきことは、これらの同じネットワークアプローチが、人間の社会的相互作用を説明し、伝染病、喫煙、肥満などの問題がどのように人間集団内でクラスター化し、広がるのかを理解するのに関連しているということです。
ネットワークがどのように動作するかを理解するには、システム構造 (ネットワークがどのように組織され、相互に接続されているか)、システムダイナミクス (時間の経過とともにどのように変化し、摂動にどのように反応するか)、制御方法 (システムがどのように自身を調整するか) などのいくつかのパラメーターを説明することが重要です。または他のシステムによって規制されている)、および全体的な設計方法(どのように変更され、成長するか)。ネットワーク理論の脳システムへの応用はまだ比較的新しいですが、精神医学および精神神経疾患について多くの洞察を提供しています。
ネットワーク組織に関する初期の研究は、1950 年代に Paul Erdos らによって開発された数学的原理に基づいており、「通常の」ネットワークの構造を扱いました。このようなネットワークでは、各ノードには他のノードとほぼ同じ数の接続があります。 「ノード」とは、ネットワーク内の個々の接続ポイントです。たとえば、ソーシャル ネットワーク内の 1 人の人物、シナプス ネットワーク内のニューロン、高次の神経システム内の脳領域などです。通常のネットワーク モデルは、各主要都市にほぼ同数の主要道路が出入りする米国の高速道路システムのような一部のネットワークを記述するのに適しています。問題は、生物学的および人間のソーシャル ネットワークがより複雑であり、ノードが持つ接続の数、接続がどのように組織されるか、接続の強さに関して大きく異なる可能性があることです。
ネットワーク科学は、1960 年代と 1970 年代に社会科学者によって進歩されました。人間のネットワークには、通常のネットワークと同様に高度なローカル クラスタリングがあり、密接な関係にある個人を反映しているだけでなく、効果的な情報伝達を可能にするクラスタ間のショートカットも存在することがわかりました。これはスタンリー・ミルグラムによって概説された「スモールワールド」の接続性の本質であり、人間が互いに高度な関連性を持っていることを示しています。これは「6 次の隔たり」として有名で、すべての人間が社会的絆を通じて互いにつながっていることを示す現象です。私たちの誰も、地球の最も辺鄙な地域の人々から約 6 つ以上離れたところにいません。したがって、友人の友人の友人、あなたの個人的な友人などを通じて、地球上のかなり離れた地域にあなたを繋ぐことができます。
————————図 3-1
スモールワールドネットワーク
スケールフリーネットワーク
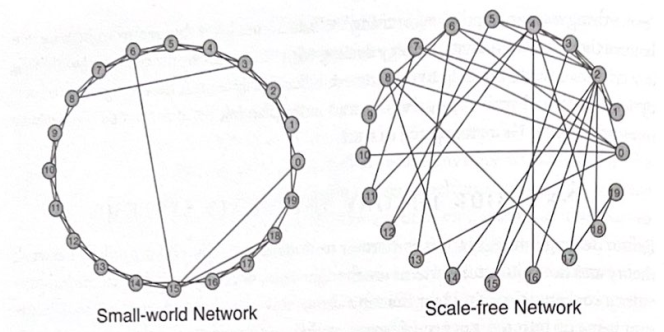
図 3-1 スモールワールド ネットワークとスケールフリー ネットワーク。左側のグラフは、通常のネットワーク内にいくつかの長距離接続が介在するスモールワールド ネットワークの接続を示しています。右のグラフは、いくつかの長距離接続とともに高度に接続されたノードがいくつかあるスケールフリー ネットワークを示しています。 (モデル提供: セントルイスのワシントン大学、ラリー アイゼンマン)
————————-
ダンカン・ワッツとスティーブン・ストロガッツは 1990 年代に、回虫である線虫の神経系における細胞の接続性など、特徴が明確ないくつかの現実世界のネットワークを分析することで、これらの概念をさらに追求しました。研究者らは、これらのネットワークの接続パターンが「スモールワールド」トポロジーに従っていることを発見しました。このトポロジーでは、少数の長距離接続を伴う高度なクラスター化が行われ、特定のクラスターが他のクラスターに大きな影響を与えることができました。このトポロジは、完全に規則的な接続と完全にランダムな接続の中間に位置します。このような「スモールワールド」ネットワークは、それらが呼ばれるように、その経路長 (任意の 2 つのノード間の距離の尺度) とクラスタリング係数 (ノードのローカルな近傍の関連性または接続性の尺度) によって記述されます。 。 「スモールワールド」現象は、ローカル レベルで非常に高度にクラスター化されているネットワーク内で、いくつかの長距離ショートカットが原因でパス長が大幅に減少した場合に発生します (図 3-1)。重要なのは、ネットワーク機能に大きな影響を与えるために、長距離接続がそれほど強力である必要はないということです。マーク・グラノヴェッターは 1970 年代に、人間がタスクを実行するためにソーシャル ネットワークをどのように使用するかに関する研究の中で、このことを説得力を持って説明しました。たとえば、新しい仕事を探すとき、最も助けとなるのは、自分の社会的領域のより周縁にいる人々との弱いつながりであるのが一般的であり、その主な理由は、これらのつながりが親しい同僚のつながりとは異なり、補完するためである。グラノヴェッターはこれを「弱い絆の強さ」と呼びましたが、神経科学の現在の研究では、この概念が人間の社会的ネットワークと同じくらい脳の機能に関連している可能性があることが示されています。
Albert-Laszlo Barabesiらは、生物学的ネットワークおよび社会的ネットワークにおけるつながりの分布を調べることでこの研究を拡張した。彼らは、実際のネットワークには「スケールフリー」タイプの接続があると提案しました。ここで、接続の分布は「べき乗則」に従い、システム内のほとんどのノードには少数の接続しかありませんが、ネットワーク内の少数のノードには多数の接続があります。したがって、ノードが持つ接続数の分布は、ネットワーク内のノード数に応じて急速に減少する関数であり、「スケール」するための尺度を作成するためのネットワーク内の真の「平均」接続数は存在しません。ネットワーク (したがって「スケールフリー」と呼ばれます)。これは、「ベル型曲線」で表される接続の「正規分布」とは区別されます。ここでは、大部分のノードがほぼ同じ数の接続を持っているため、真の「平均」接続数が存在することになります。
重要なのは、スケールフリー トポロジは自己組織化システムの特徴であり、その結果、接続のハブが存在するネットワークが形成されます (米国の航空路線の地図とよく似ており、ニューヨークやロサンゼルスなどのいくつかの都市では、高度な接続が行われます)。トピカやオマハなど、他のほとんどの路線は発着するルートが数本しかありません) (図 3-1 を参照)。このタイプの分散は「80-20 の法則」と呼ばれることもあります。つまり、20% のノードが 80% の接続を持っています。これは、人口内の富の分布、または書かれたテキスト内の単語の分布を記述するために使用される場合、それぞれパレートの法則 (経済学) またはジップの法則 (言語学) とも呼ばれます。スケールフリーの接続分布は、米国の送電網、インターネット、線虫の神経系など、多くの生物学的および社会システムで見られます。また、ネットワークのダイナミクスにも大きな影響を及ぼし、ネットワークがどのように発展し、成長するかを予測する可能性があります。たとえば、ネットワークの成長は、少なくとも部分的には、高度に接続された既存のノードに新しいノードが優先的に接続されることに起因すると考えられています。事実上、バラベシ氏の言葉を借りれば「金持ちはさらに金持ちになる」のだ。このようなシステムは、個々のノードに対する攻撃によってネットワーク機能が中断されることはほとんどないため、非常に堅牢です。これは、ほとんどのノードには少数の接続しかなく、ネットワーク機能にとって重要ではないためです。対照的に、高度に接続された少数のノードが損傷すると、ネットワークが大幅に混乱する可能性があります。したがって、スケールフリー ネットワークは、ほとんどの個々のノードへの損傷に対して非常に耐性がありますが、十分な数の主要ノードが中断されると、連鎖的な障害が発生する可能性があります。
スモールワールドおよびスケールフリーのネットワークに関するこの研究は、システム神経科学に関連しています。これらのタイプの接続性は、ICN が脳内でどのように組織化されているかを理解するために非常に重要であると思われ、また、特定の脳構造が複数のシステムに現れ、「すべてが接続されている」という重要だがもどかしい考えにつながる理由を説明するのにも重要である可能性があります。すべてに。」現在のデータは、脳が階層ではなく一連の閉ループである回路に組織化されているという考えを裏付けています。これらの脳ネットワークには、効率的な情報の流れにとって重要なスモールワールド特性があります。スケールフリー トポロジが関係しているかどうかはあまり明らかではありませんが、一部の脳領域は他の領域よりも強く接続されており、べき乗則接続などのハブのような特性を持っています。私たちは議論の中でスモールワールドとスケールフリーの両方の視点を使用しますが、これが進化する科学分野であることは容易に認めます。これらの概念は、なぜ脳がこれほど堅牢であるのか、また、まれな例外を除いてなぜ脳がコンピューターのように「クラッシュ」しないのかを理解するためにも重要であると私たちは考えています。しかし興味深いことに、最近の研究では、相互に依存する一連のネットワークのうちの 1 つのネットワーク内のノードの機能不全が、損傷を受けていないネットワークにどのような悪影響を及ぼす可能性があるかを調査しています。これらの観察は、なぜ精神疾患が複数の神経系の機能不全を引き起こすのかを理解するのに関連している可能性があります。最近の研究では、「ストレス」がネットワークの開発とトポロジにどのような影響を与えるかについても調査されており、その結果、システムに対するより穏やかなストレスが、より堅牢で自立する手段として、最初はランダムなネットワークをスケールフリー接続に向けて推進するのに役立つことを示唆しています。残念ながら、ストレスが大きくなり持続すると、高度に接続されたハブが損傷し、接続の崩壊やシステムの故障が発生する可能性があります。ストレスとネットワーク機能のテーマには後の章で戻ります。また、ニューロンネットワークでは、さまざまな形態のシナプス伝達 (興奮性と抑制性) がネットワークのトポロジーと活動に大きな影響を与える可能性があります。図 3-2 は、培養海馬ニューロンのネットワークにおける高速 GABA 媒介阻害の操作の影響を強調する実験の結果を示しています。
————————図 3-2
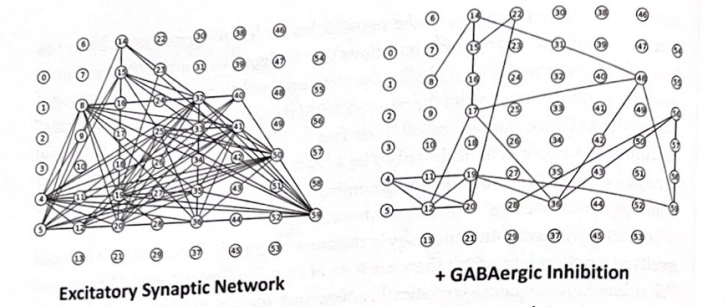
図 3-2 ニューラル ネットワークに対する抑制の影響。この図は、マルチ電極アレイ (MEA) 記録を使用した、ラット海馬ニューロンの培養におけるニューロン発火の相関関係を示しています。図中の小さい番号の丸は記録電極を表す。グラフ内の線は、調整された (相関した) 方法で発火するニューロンを示しています。左側の図に示すように、いくつかの「ノード」は高度に接続されています (例: #59)。 GABA作動性阻害がネットワークに追加されると、右の図に示すように、相関発火が減少し、接続パターンが劇的に変化します。 (データ提供: セントルイスのワシントン大学、ラリー アイゼンマン)
————————
脳の組織
脳システムが精神疾患にどのように寄与しているかを議論するには、脳がどのように組織されているかについて「全体像」を理解することが重要です。この議論のために、私たちは再びジョゼフ・ルドゥーと彼の著書『シナプス・セルフ』から借用します。 LeDoux は、精神科医が理解するために重要ないくつかの基本概念について説明しました。これらの概念の一部は自明であり、些細なことのように思えるかもしれませんが、ICNS と精神科の症状や病気における ICNS の役割についての後の議論に深い意味を持ちます。
高いレベルから見ると、脳は並列処理システムの集合として組織されています。これらの並行システムはすべて、同じ情報、つまり個々の人間の世界を認識し、それに基づいて動作します。これは些細なことのように思えるかもしれませんが、情報を同時に処理する並列システムが多数存在するため、ある脳システムと別の脳システムで情報が処理される方法と時期の不一致は、「現実」に大きな歪みをもたらす可能性があります。これは明らかに、第 1 章で説明した切断症候群の根本的な欠陥です。脳内にいくつの処理システムが存在するかについては議論がありますが、その数は比較的多く、サブシステムの分類方法にもよりますが、少なくとも 15 です。たとえば、私たちは 5 つの主要な感覚 (視覚、聴覚、味覚、嗅覚、触覚) のそれぞれから個別の入力を受け取ります。これに、バランス、覚醒、注意、言語、記憶、空間情報、感情、報酬、神経内分泌制御などのシステムが追加されます。出力を生成するためのシステムも複数あります (運動システムや言語出力システムなど)。最も単純なレベルでは、一次感覚システムは世界についての基本的な認識を提供します。ただし、これらの「知覚」は実際には複雑な経験であり、通常は 1 つの特定の様式における 1 つの感覚入力のみを反映しているわけではないことを認識することが重要です。少なくとも、経験には複数のシステムが関与していることが多く、実際の認識は、感情や記憶などの内部情報と外部情報の両方によって味付けされます。このため、単純で純粋な知覚というものが存在するかどうかについては議論の余地があります。
これらのさまざまな並行システム間で活動を柔軟に調整して、現実の一貫した画像を生成する方法は、脳にとって大きな課題です。脳がそれを達成する方法の 1 つは、並列システム内および並列システム間のシナプスの構造と機能が活動に依存して変化することであり、このプロセスは広く「可塑性」と呼ばれます。脳の可塑性の主な要因は、入ってくる情報の時間的相関関係であると考えられます。つまり、異なるシステムが同時に発生していると認識する情報は、それらのシステムが協調して動作するかどうかに大きな影響を与えます。グルタミン酸を神経伝達物質として使用する脳の高速興奮性シナプスは、時間的に密接に相関して発生する活動を検出するのに非常に優れています。情報を送信および受信するニューロン (それぞれシナプス前ニューロンとシナプス後ニューロンと呼ばれる) の同時活動は、それらのニューロン間の接続の強化をもたらします。このプロセスは、特定の種類の記憶の基礎。多くの脳シナプスがヘブ可塑性を起こすことが現在では明らかであり、この過程が「一緒に発火するニューロンは一緒に配線する」という格言の基礎となっている。この種の可塑性により、脳領域は相互の偶然の経験に基づいて互いに結びつく能力を持っています。対照的に、一時的に不調和な活動は、ニューロンと脳システムの間の接続の弱体化につながります(反ヘビアン可塑性と呼ばれます)。これら両方の形態の脳可塑性の根底にあるメカニズムは、記憶処理の根底にある分子生物学および構造生物学に関する重要な洞察をもたらし、多くの主要な精神神経疾患における機能不全を矯正するための潜在的な標的を提供しています。これらのメカニズムと機会のいくつかについては、本書の後半で取り上げます。
可塑性に起因する変化に加えて、脳はシステム内およびシステム間の活動を調整するために、いくつかの種類の神経伝達物質および神経調節物質も使用します。神経伝達物質はいくつかの方法で分類できます。たとえば、特定の伝達物質(グルタミン酸やアセチルコリンなど)は、ニューロンを脱分極させ、ニューロン内の高速細胞内コミュニケーションとニューロン間の高速細胞間コミュニケーションを促進する電気信号である活動電位の発火を容易にするため、「興奮性」です。他の伝達物質(GABA やグリシンなど)は、ニューロンが活動電位を発火するのを困難にするため、「抑制性」です。他のスキームでは、化学構造 (生体アミン、アミノ酸、ペプチドなど) に従って送信物質を分類します。
脳内の活動の調整を理解するには、Bertil Hille や他の生理学者が説明しているように、神経伝達物質が機能する時間経過に基づいて神経伝達物質について考えることも役立ちます。この方式では、送信機は「速い」または「遅い」と言われます。高速伝達物質 (グルタミン酸や GABA など) は、シナプス前ニューロンから高濃度で放出され、受容体上で短時間 (ミリ秒の時間スケール) 急速に作用し、シナプス後 (受信) ニューロンの細胞膜にイオン チャネルを開きます。イオンチャネルは通常、伝達物質(場合によっては細胞膜を横切る電気勾配の変化、またはその両方)に応答して開閉するマルチサブユニットタンパク質の細孔です。これらの細孔はイオンを細胞に出入りさせ、その過程で細胞膜全体の電気勾配を変化させます。グルタミン酸は、正に荷電したイオン (ナトリウム、カリウム、カルシウム) が膜を通過できるようにするカチオンチャネルを開きます。これにより、脱分極 (励起) が生じます。 GABA は、マイナスに帯電したイオン (塩化物やその他のアニオン) が膜を通過できるようにするアニオン チャネルに作用します。これは通常、常にではありませんが、過分極 (阻害) を引き起こします。一般に、伝達物質が興奮性であるか抑制性であるかを定義するのは、膜電位に対する影響です。高速送信機は脳内で瞬間瞬間の動作を提供し、事実上、多くの「利益」をもたらします。表 3-1 は、高速および低速の送信機システムがどのように構成されているかをまとめたものです。
対照的に、低速伝達物質は通常、シナプス領域に低濃度で存在し、より拡散的に作用し、脳領域内および脳領域全体で活動を組織化し調整する方法を提供します。これらの伝達物質は、化学的なセカンドメッセンジャーシステムに結合する別の種類の受容体にも作用します。これらの遅い伝達物質の影響は、多くの場合、伝達物質の受容体への結合と、受容体を細胞内メッセンジャーに結合させる連結タンパク質(グアニンヌクレオチドが関与しているため、Gタンパク質と呼ばれる)の活性化を含む多段階のプロセスで発生します。システム。 G タンパク質は、受容細胞内でさらなる化学変化を引き起こす特定の酵素の活性化を助けます。たとえば、アデニル酸シクラーゼの活性化とサイクリック AMP の生成、またはホスホリパーゼ C の活性化とイノシトール三リン酸 (IP3) とジアシルグリセロールの生成です。重要なのは、これらの遅い伝達物質は、数秒から数分、あるいはそれ以上持続する効果を生み出し、その結果、細胞の機能と興奮性に対してより長く持続する効果をもたらすことです。これらの伝達物質が開始する化学事象は、細胞内で化学事象をさらにカスケードおよび増幅させ、最終的には細胞核内の遺伝子の発現を促進または阻害する可能性があります。遺伝子発現は、受容体の合成、細胞増殖、および細胞コミュニケーションに長期的な影響を与えるその他の構造変化を指示することができます。
————————表 3-1

表 3-1 高速神経伝達物質と低速神経伝達物質
グルタミン酸/GABA (速効性)
高速信号 (<100 ms 持続時間)
小胞は活性ゾーンにドッキングし、
プライミングされています
放出: 局所的に高濃度の Ca2+ が必要
生体アミン (遅い)
低速で持続的な信号 (秒)
小胞は活性ゾーンやその他の場所から放出される可能性があります
放出: 持続的な低 Ca2+ 上昇
急速な除去 (拡散、トランスポーター、マイクロ秒) 遅い除去 (トランスポーター、秒)
ポイントツーポイント伝送
シナプス、離散
受容体: 低親和性リガンド依存性イオン
チャネル (LGIC)
スピルオーバー(パラクリン効果) 体積透過
受容体: 高親和性 G タンパク質共役型 (GPCR)
————————
1つのシナプス前細胞が、異なる脳領域での送信機の放出を含め、伸びて分岐する軸索に沿った複数の場所で同時にその送信機を放出する可能性があるため、これらの遅い伝達物質は拡散的に作用する。遅い送信機の影響下で、受信セルは、高速な送信機からの着信メッセージに多かれ少なかれ応答する状態になります。これは、シナプス全体および脳の領域全体の活動を調整する機会を提供します。実際、これらの遅い送信機は神経細胞や神経領域の「調子を設定」し、より速い入力が影響を与える背景を確立します。遅い伝達物質の例には、向精神薬(ドーパミン、セロトニン、ノルエピネフリンなど)によって変化する神経調節物質の多くが含まれます。多くの高速送信機は、自身の送信機に特有の低速受容体にも作用して、自身の高速活動を調節するさらに別のソースを提供することに注意することが重要です。たとえば、グルタミン酸、GABA、およびアセチルコリンは、それらの高速リガンド依存性イオンチャネルに加えて、「代謝指向性」または G タンパク質共役受容体 (GPCR) にも作用します。 「リガンド」という用語は、受容体に結合する化学物質を指し、「ゲーティング」という用語は、イオンチャネルを開いたり閉じたりする薬剤の能力を指す用語です。図 3-3 は、神経伝達物質受容体の構造を示す図です。
脳の領域全体で活動が調整されると、出力を駆動するだけでなく高次の処理にも役立つように、何らかの方法で統合する必要があります。接続性の高い領域、特に新皮質の領域は、処理に独自の計算次元を追加しながら、この情報をまとめる上で重要な役割を果たしているようです。これらの領域は、LeDoux の用語では「収束ゾーン」と呼ばれ、海馬傍、後頭頂皮質、および前頭前皮質が含まれます。これらの領域はマルチモーダル入力を受け取り、前述のスケールフリー ネットワークの高度に接続されたノードのようなものです。これらの接続性の高い領域により、入力をより複雑な表現に合成できます。ルドゥー氏は、これらの領域は認知合成と高次処理を反映して、「認識」を「概念」に変えることができると説明しています。重要なのは、新皮質の非常に高い接続性を有する領域は、脳の下位中枢にも影響を与える可能性があり、直接的な感覚経験からシナプス的に除去された高レベルの抽象的思考が、「上部」を介して独自のその後の入力に影響を与えることができる、領域全体にわたる調整のさらに別の形式を提供する可能性があります。 -ダウン処理」、フィードバック (さらには意識的) 制御の一種。この種の認知制御は人間 (または少なくとも高等霊長類) に特有のものであり、抽象的な思考が感覚、感情、動機、記憶に影響を与える方法です。
————————図 3-3
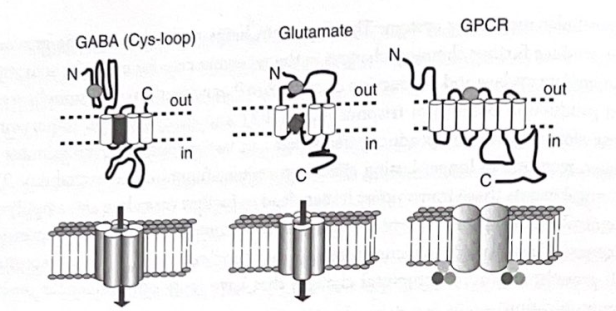
図 3-3 3 つのクラスの神経伝達物質受容体。これらの図は、神経伝達物質受容体の 3 つの主要なクラスの提案された構造を示しています: イオンチャネル性 GABA-A 受容体 (ニコチン性アセチルコリン受容体およびグリシン受容体とともに Cys ループファミリーに属する)、イオンチャネル性グルタミン酸受容体 (AMPA、カイニン酸、および NMDA を含む)タイプ受容体)、および G タンパク質共役受容体(GPCR)の広範なファミリー。下の図は、イオンチャネル受容体の場合に複数のサブユニットがイオンチャネル機能にどのように寄与すると考えられるか、また二量体化と G タンパク質への結合が GPCR 活性にどのように寄与すると考えられるかを示しています。 (図は、セントルイスのワシントン大学の Steve Mennerick 氏のご厚意によるものです。)
————————
収束ゾーンは、新しい記憶の形成に重要な海馬と呼ばれる脳の領域にも入力を提供します。海馬は、新皮質の 6 層構造とは異なる 3 層構造をしているため、「古皮質」と呼ばれる進化的に古い脳の部分です。この領域は、場所と時間に関するデータを提供する連合皮質からの高レベルの入力、扁桃体やその他の領域からの感情入力、私たち自身の身体の内部状態や脳機能に関する情報など、幅広い情報を処理します。ヘビアン可塑性を利用して、海馬はこれらの情報をまとめて新しい記憶を形成します。実際、この領域は、物事を思い出すことを可能にする短期記憶痕跡を生成しながら、新規性、関連性、および文脈を処理するのに役立ちます。次に、海馬は処理されたデータを連合皮質および他の脳領域に送り返し、そこでより永続的な記憶痕跡が生成されます。海馬は新しい情報と以前に処理された情報の両方を扱うため、古い記憶を取り出して新しいものに変えることができます。これにより、何をどのように記憶するかについて大きな柔軟性が得られるだけでなく、記憶処理が過去と現在の状態の両方に大きく影響される可能性もあります。また、私たちが自分自身を未来に投影する能力は、主に記憶に基づいています。したがって、海馬は将来の取り組みを計画するためにも重要です。海馬は、その高度な接続性とその入力の性質により、ルドゥーが「超収束ゾーン」と呼ぶものを表しており、大うつ病、双極性障害、統合失調症、およびストレス関連障害。これは、新しい宣言的記憶の形成にとって重要な領域です。
スケールフリー ネットワークで主要なノードの損失が連鎖的なネットワーク障害を引き起こす可能性があるのと同様に、何らかのメカニズムによる海馬機能の損失は、認知機能に壊滅的な影響をもたらします。
これらの基本原則は、高レベルの情報を処理し、精神疾患の症状を生成する際に重要な役割を果たす脳ネットワークである ICN について考えるための準備を整えます。 ICN は主に、機能的磁気共鳴画像法 (fMRI) 研究で観察される相関のある BOLD (血中酸素濃度依存性) 信号に基づいて特定されています。 BOLD 信号は、脳領域の局所的な活動から生じる酸素化ヘモグロビンと脱酸素化ヘモグロビンの比率を測定します (ヘモグロビンの磁気特性の変化によって検出されます)。したがって、信号は脳血流と酸素消費量の局所的な変化を伴う血行力学的反応です。その生理学的相関関係は不明ですが、脳領域の統合された神経活動の電気信号である局所電場電位 (LFP) を反映していると考えられています。 BOLD を使用した研究により、特定の脳領域の活動におけるコヒーレントな低周波変動が 1 秒あたり約 0.01 ~ 0.1 サイクル (ヘルツ [Hz]) の速度で発生していることが明らかになりました。したがって、これらの機能の相関関係は、活動電位の発火と高速なシナプス伝達に関連するミリ秒の時間枠よりもはるかに遅い、数十秒の時間枠で発生します。これらのゆっくりと変動する信号は、脳のリンクされた領域間で高度に相関しており、活動の接続マップの生成を可能にします。これについて簡単に考えると、接続された脳領域は、同時の活動で一緒に「ハミング」しているように見え、この「ハミング」(およびそれに関連する接続性)が脳内の高次の処理を駆動している可能性があります。この遅い協調的な活動を引き起こすものはまだ明らかではありませんが、ニューロンとグリアによって提供される神経調節物質が寄与している可能性があります。神経伝達物質が脳ネットワークと精神処理にどのように寄与するかについての詳細はまだ不明ですが、現在の考え方の概要を表 3-2 に示します。
多数の主要な ICN が記載されており、将来さらに多くの ICN が発見されることは間違いありません。第 2 章では、脳が他のタスクに集中していないときに自己関連情報を処理すると思われるデフォルト ネットワークと、大脳辺縁系ネットワークに関与する感情顕著性システムについて説明しました。現在認識されている他の ICN には、空間注意を制御する分散神経システム (おそらく非優位半球の少なくとも 2 つの前頭頭頂ネットワークからなる)、言語 (優位半球のペリシルビウス ネットワーク)、顔と物体の認識 (下側頭ネットワーク)、意思決定 (前頭前実行ネットワーク)、および認知制御(少なくとも 2 つの前頭頭頂ネットワーク)。これらの ICN のいくつかと、それらが心や精神疾患にどのように関与しているかについては、本書の後半で改めて説明します。表 3-3 は、現在特定されているいくつかの ICN の概要と、それらが人間の心の三者機能に寄与するものとしてどのように概念化されるかを示しています。構造的相関関係に興味がある方は、付録および図 2-1 および 2-2 を参照してください。
————————表 3-2

表 3-2 高速伝達物質と低速伝達物質: 行動における役割 グルタミン酸/GABA (高速)
ミリ秒思考 自発的行動
記憶のエンコードと取得
抗けいれん薬、神経保護薬、抗不安薬、抗うつ薬の標的
生体アミン (遅い)
覚醒
注意
気分
褒美
抗うつ薬、抗不安薬、降圧薬の標的
————————表 3-3

表 3-3 マインドと ICNS
認知(思考)
作業記憶ネットワーク (背外側前頭前野-頭頂葉) 注意/方向転換ネットワーク (前頭頭頂葉)
前頭前幹部ネットワーク
空間ナビゲーションネットワーク(前頭頭頂)
言語ネットワーク (ペリシルビア語)
顔および物体認識ネットワーク (下側頭)
感情(意味)
拡張された扁桃体処理ネットワーク
感情顕著性ネットワーク
明示的記憶ネットワーク (辺縁系)
モチベーション(目標)
腹側被蓋野-腹側線条体モチベーションネットワーク 報酬処理ネットワーク
————————
感情とモチベーションに関係する脳システムは、この組織図にどのように適合するのでしょうか?海馬と同様、これらのシステムは進化的に古く、環境を迅速に評価し、その環境をうまくナビゲートする方法を提供することで人間の生存を助けるために進化したと考えられます。それらは独立して動作することができ、効果的に機能するために新皮質や海馬を必要としません。これらのシステムは通常、皮質と海馬と連携して機能しますが、トリガーされると制御を引き継ぐ能力を持っています。怒りの感情を経験したことのある人なら誰でも、これを証明できます。激怒は明らかに理性を無視し、攻撃的な行動を引き起こす可能性があり、それが個人にとって有益なこともありますが、皮質が再び動き始めて何が起こったのかを評価すると後悔することがよくあります。簡単に言えば、人は激しい一次感情を経験するために大脳皮質を必要としません。これらのシステムは人生の重要な部分であり、その活動は良いことも悪いことも含めて、私たちの経験や記憶に大きな影響を与えます。感情的および動機付けのネットワークは、高速送信機と低速送信機の組み合わせを使用して、その接続性を利用して、新皮質の高次処理センターの活動を含む、広範囲に分散した脳領域と機能の調整を支援します。
脳機能の「全体像」の原理
すぐにニューラル システムと ICN に戻ります。ここまでの説明は、全体的な脳の組織に関する背景と、精神科の症状や症候群を考えるときに脳の組織がどのように概念化されるかを提供することを目的としています。この部分の議論は、脳機能のいくつかの追加原理を説明することで終了します。ここでも、脳システムがどのように組織され、どのように機能し、病気でどのように機能不全に陥るかを考えるのに役立つ高レベルの概念に焦点を当てます。
この議論では、リード モンタギューが著書『なぜこの本を選ぶのか?』で説得力を持って説明されている原則のいくつかを借用します。まず、脳は非常に効率的な処理装置です。これは通常の意味での「コンピューター」ではありませんが、脳が現代のコンピューターとどのように比較できるかを考えることが役立つ場合があります。このような分析では、脳は最も単純なパーソナルコンピュータに比べて遅いにもかかわらず、高度な電子機器が困難を伴うタスクを容易に実行できることが明らかです。顔認識と複雑なパターン補完は 2 つの例です。高速なコンピューターは多くの電力 (エネルギー) を必要とし、無駄な努力によって大量の熱を発生するため、脳の速度が遅いことは実際には少しありがたいことです。実際、脳はエネルギーの観点から非常に要求が高く、機能を維持するには心拍出量の約 20% ~ 25% を必要としますが、その量は人間の体重の約 2% ~ 3% にすぎず、どのような基準から見ても驚くべきエネルギー需要です。しかし、おそらくさらに驚くべきことは、極端な状況でない限り、脳は過熱しないという事実です。部分的には、コンピューターに比べて速度が遅いため、熱の放散に役立ちます。遅いデバイスは、高速なデバイスほど発熱しません。精神科医にとって、脳は多くのエネルギーを必要とし、エネルギーが不足したり疲労したりするとうまく機能しないことを認識することが重要です。これはおそらく、人の脳がトップダウンの処理を行っているときに観察するのが最も簡単です。たとえば、感情などのより原始的な機能をオーバーライドして制御するために高次の認知が使用される場合です。疲労と休息の欠如は、感情のコントロールに悪いことで知られており、多くの考え抜かれた行動の一因となる可能性があります。
第二に、脳はエネルギーと労力を節約するためにショートカットを使用するため、効率的な処理装置です。曖昧または不確実な刺激に直面すると、脳は状況を一貫性のあるものにするために、欠けている詳細を補うことがよくあります。これは、曖昧な感覚入力 (ゲシュタルト図など) またはより複雑な状況 (社交的な出会いなど) で発生する可能性があります。モンタギューが説明したように、脳の計算は不正確であり、スペースとエネルギーを節約するために脳は多くの情報を圧縮する必要があります。これは計算効率にとっては素晴らしいことですが、脳が必ずしも「真実を生成する装置」ではないことを意味します。内部および外部の現実に対処するためのスキームやパターンを作成および使用しますが、このアプローチは情報の処理方法と解釈方法に偏りをもたらします。もう一度、感情が状況を迅速に把握するのにどれほど効率的であるか、しかし感情がどれほど間違っている可能性があるかを考えてください。
これは、3 番目の一般原則につながります。それは、脳が人の「現実」を創造するということです。精神科医は、幅広い精神疾患にわたってこの事実に日常的に取り組んでいます。脳が「現実」の創造者として機能していることは、精神病性障害において最も明らかかもしれない。精神障害では、誤った認識、誤解、単なる論理的誤りが、世界の仕組みについての固定的で奇妙で誤った信念につながる可能性がある。しかし、精神病患者にとって、それは彼らが認識する現実です。同様に、認知行動療法 (CBT) などの特定の精神療法は、患者が世界と自分の人生についての誤った認識を理解し、修正できるように設計されています。繰り返しになりますが、うつ病患者にとって、世界に対する否定的な偏見は彼らの現実ですが、それは必ずしも中立的な観察者の現実ではありません。脳が現実を創造するという考えは精神疾患の場合に最も理解しやすいかもしれませんが、この現象は病気の人に特有のものではありません。実際、それは私たち全員の中に起こります。ひとたび解釈と信念の領域に足を踏み入れると、多くの人間が現実について興味深い、しかし必ずしも事実や論理的な解釈をしているわけではないことが明らかになります。たとえば、政治と宗教について考えてみましょう。人間はあらゆる種類の奇妙なことを信じており、状況や結果に関係なく、しばしば極度の信念を持ってそれらの信念を保持します。堅固な意見と妄想との違いは、定性的なものではなく量的な区別である可能性があり、一部の観察者は精神病性の「妄想」と非精神病性の「妄想」を区別しています。
正常な脳が現実を創造し、間違いを犯すとしたら、精神疾患を患う人とそうでない人を概念的に分けるものは何でしょうか?違いの 1 つは、私たちの脳が日常的に犯す「間違い」を修正するのに十分な認知的および感情的な柔軟性を備えているかどうかにあるかもしれません。すべての人間は機嫌が悪くなったり、考えが悪くなったり、社会的交流が悪くなりますが、深刻な精神疾患を発症する人は少数です。ほとんどの人は、意識的または無意識的に、誤解や気分の変化を修正する方法を見つけます。モンタギューは、誤り訂正は脳の情報処理にとって重要であり、部分的には、脳がその計算と出力を導くために目標(動機)と期待を使用する戦略の結果であると主張した。目標と期待は内部で生成され、新しい経験に合わせて変更できます。私たちの脳は、私たちが認識し経験する「現実」の結果を、内部の期待や目標と比較し、期待が満たされない場合にエラー信号を生成します。神経伝達物質であるドーパミンはこのプロセスにおいて重要な役割を果たしているようで、ドーパミンを生成する中脳ニューロンの発火の相対的な変化は、結果が期待どおりかどうかを反映しているようです。行動と思考は、認識された不一致に対処するためにバイアスがかかります。後者の計算には、ドーパミン-腹側線条体系内の重要なノードに接続されている吻側前帯状皮質の領域が関与しているようです。これにより、脳がその「仮想世界」を更新および修正し、より効率的に目標に向けた行動を実現するために期待を現実と一致させようとする一種の「強化学習」が確立されます。この重要な誤り訂正プロセスにおけるドーパミンの役割は、ドーパミン媒介伝達の変化が主要な神経精神障害において重要な役割を果たしているように見える主な理由の 1 つである可能性があります。
最後に、強化学習、および学習一般は脳の可塑性を反映しています。人間の脳はすべて「可塑性」があり、経験に依存した学習と記憶が可能です。我々は以前、これをヘブ型シナプス可塑性として説明し、神経系機能における組織化原理としてのその役割を議論しました。脳のこの「可塑性」特性は、どれだけ強調してもしすぎることはありません。もし脳が可塑性でなかったら、精神病患者には希望が持てないでしょう。私たちは、精神医学で使用されるすべての治療法は脳の可塑性を介して機能し、神経回路が経験に基づいて適応し再構成できるという事実が、病気の最適な転帰を達成するために重要であると主張します。したがって、私たちの見解では、精神科医は脳の可塑性を理解し、広く概念化された可塑性を治療目的に利用する専門家にならなければなりません。
ICNS の基本原則
ICN の識別方法を含め、ICN に関する一般的な概念を説明するために、すでに述べた ICN、つまりデフォルト システムに関する理解の進化を振り返ります。ここではデフォルト システムに焦点を当てますが、概要を説明した原則の多くは他の ICN にも関係することを理解することが重要です。第 2 章 (表 2-1 および図 2-1) では、このネットワークの主要な構造と機能の概要を示しました。ここでは、デフォルト システムのような ICN が機能的神経画像研究に基づいてどのように特定されるかに焦点を当てたいと思います。
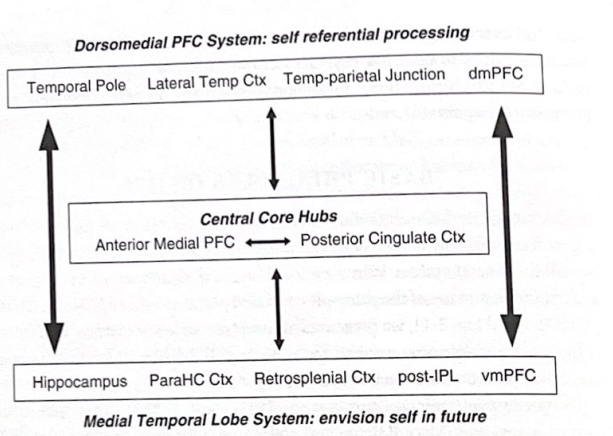
Marc Raichleらは、陽電子放射断層撮影法(PET)とBOLD機能的磁気共鳴画像法を用いて、比較的長い(10秒以上)間隔で検査すると、特定の脳領域が高度に相関した活動を示すことを実証した。人が休息していて特定のタスクに取り組んでいないときに、脳はこの相関した活動を示します。この「休息」活動が、広範囲に分散した特定の脳領域間で高度に相関しているという発見は、驚くべきものであり、新しいものでした。この休止本来の活動に関与する領域は、「デフォルト モード ネットワーク」を定義するようになりました。人がタスクに集中し、特定の何かに積極的に注意を払い始めると、デフォルトのネットワークの活動が減少し、注意システムや実際のタスクの実行に関与するシステムを構成する脳領域の活動が増加します。このため、一部の認知科学者は、ニューラル ネットワークを「タスク依存」または「タスク独立」として概念化しました。実際、初期の研究では、課題に依存して活動が一貫して低下する領域が繰り返し観察されたことから、Raichle らは、課題を行っているときよりも安静時のほうが活発な重要な脳ネットワークに取り組んでいる可能性が高いと考えました。その後の分析により、これらの同じ構造間には静止状態での高い活性と一貫性が存在することが実証されました。デフォルトのネットワークを構成する分散した脳領域の相関変動は、人が起きているときも、休んでいるときも、さらには麻酔を受けているときも発生することが判明しました。このシステムの一貫した活動は浅い睡眠でも持続しますが、深い(徐波)睡眠中は前頭前皮質の関与が減少し、一部の領域が切り離され、おそらく領域間の興奮と抑制の変化を反映しています。後者の発見は、デフォルトのネットワークが意識的な認識の経験に寄与している可能性があり、ネットワーク接続が動的で状態に依存する可能性があることを示唆しています。図 3-4 は、デフォルト ネットワークを構成するいくつかの主要な脳領域の図を示しています。
重要なのは、このネットワーク内のほとんどの場所は実際にはそれ自体がハブであるにもかかわらず、デフォルトの ICN 内の一部の領域は他の領域よりも機能的に接続されているということです。接続の程度の違いを反映して、メインのデフォルト ICN 内にサブシステムが存在する場合もあります。領域が機能的にどのように接続されているかを判断するために、研究者は対象となる特定の脳構造の活動を調べ、その構造の活動を他の脳領域の活動と経時的に比較します。このような分析で主に関心のある領域は、「シード」領域と呼ばれます。たとえば、脳梁の後ろにある新皮質の一部である後帯状皮質 (PCC) は、デフォルト システムの重要なハブ領域です (脳領域のマップについては付録を参照)。それがデフォルトシステムの他の領域および他の脳システムにどのように接続されているかを判断するために、PCCを中心として機能画像データが検査されます。接続領域は、PCC の変化と正の相関がある活動に基づいて特定されます。脳の他の領域はシード領域と同期せずに共鳴し、「逆相関」している可能性さえあり、それらの領域の活動が逆方向に変化することを示しています。同様の分析が脳内の他のシード領域でも行われ、これによりそれらの固有の機能的接続性を決定することができます。これらの解析に異なるシード領域を使用することで、研究者は、対象のネットワークに関与する主要な構造を特定する際に高い統計的信頼性を得ることができます。
————————図 3-4
背内側 PFC システム: 自己参照処理 側頭極側方温度 Ctx 温度-頭頂接合部
中央コアハブ
前内側PFC
海馬
後部帯状回CTX
dmPFC
ParaHC Ctx 後脾 Ctx IPL 後 vmPFC 内側側頭葉システム: 将来の自分を思い描く
図 3-4 デフォルトのネット サブシステム。この図は、デフォルト モード ネットワークの主要な領域を示しています。現在概念化されているように、このネットワークには、自己参照情報を処理する背内側前頭前野 (dmPFC) システムと、記憶処理と未来志向の思考を扱う内側側頭葉システムが含まれているようです。これらのサブシステムは、前内側 PFC および後帯状皮質を含む中心コアのハブと相互作用します。その他の略語: ParaHC Ctx (海馬傍皮質)、post-IPL (後下頭頂小葉)、vmPFC (腹側内側前頭前皮質)。 (Andrews-Hanna 他、2010 年より改変)
————————
前述したように、ICN 内のさまざまな接続の強度は変化し、対象領域間のアクティビティの統計的相関の大きさによって決まります。たとえば、図 3-4 では、PCC と前頭葉内側前部の間の接続が強力です。これは、これら 2 つの領域の相関性が高く、他の領域よりも密接に結合しているためです。上で述べたように、特定の ICN の脳領域は、他の脳領域と強い負の相関がある場合があります。これは、他の ICN がアクティブであるときに、一部の ICN がオフになる (または少なくともアクティブが低下する) ことを示唆しています。たとえば、デフォルトの ICN の活動は、注意に関係する前頭頭頂部の ICN の活動と負の相関があります。重要なことは、精神医学にとって興味深い原則の 1 つである、ICN 内および ICN 間の接続パターンと程度の変化は、学習、生理学的状態 (睡眠段階など)、および精神神経疾患 (次の章で説明します) によって時間の経過とともに変化する可能性があることです。 )。
この議論から、特定のタスクの実行に応じた相関アクティビティの変化が、ICN がどのように定義されるかを決定する際に重要であることが明らかです。したがって、機能的神経画像と認知神経科学の科学的融合は、科学者が特定の脳 ICN と情報の計算方法を特定するのに非常に重要です。記載されている ICN の数は増加しています。これらのネットワークは、注意処理、音声処理、記憶処理、動機付けなどのさまざまな機能に関与しています。特定の脳領域は、いくつかの異なるネットワークに寄与しています。いくつかのネットワークが連携して、より高次の情報処理を可能にする可能性があります。今後数年間にわたって、ICNS の正常な機能と異常な機能に関する情報が爆発的に増え続けるでしょう。
また、特定の脳領域がいくつかの ICNS において重要なハブであることもますます明らかになってきています。これらのハブは、スモールワールドおよびスケールフリー ネットワークの説明で前述した高度に接続されたノードであり、さまざまな種類の情報を統合し、より高いレベルの認知、感情、および動機付けの処理を調整するのに役立つ領域です。これらは、LeDoux の用語では「収束ゾーン」と呼ばれます。これらのハブのいくつかはデフォルト システムの一部でもあります。これは、脳機能のさまざまな側面を調整するのにデフォルトのシステムが重要である可能性と一致しています。
要約すると、ICN は、特定の機能を達成するために共鳴 (ハム) する脳領域として特定されます。特定の ICN 内の一部のリージョンはハブであり、他のリージョンよりも接続性が高くなります。これらのハブは複数の ICN に関与する可能性があり、脳ネットワーク内および脳ネットワーク間の情報フローを調整する機会につながります。特定の機能システムのアクティビティは、別のシステムが動作しているときは特に拒否され、その逆も同様です。認知的、感情的、動機づけ的なさまざまなタスクにわたる脳活動をマッピングする研究は、ICN を特定するのに役立ち、さまざまなタスクの実行に寄与する特定の領域についての理解を深めることができます。海馬や扁桃体などの一部の領域は、行動や精神疾患に関連する複数のICNに関与しているようです。このような領域は、さまざまなシステムにおけるその役割と、それらがどのように情報を処理し、全体的な脳の接続に寄与するかという点の両方の観点から理解することが特に重要です。また、記憶処理におけるそれらの役割は、学習や新しい経験の結果としてネットワーク接続が時間の経過とともにどのように動的に変化するかという点で重要である可能性があります。
留意事項
精神障害は脳の障害です。精神科の症状や機能不全を理解するためには、精神科の症状や機能不全を脳機能の観点から概念化することがますます重要になるでしょう。
私たちは、神経科学の観点から精神障害の症状と臨床症状を理解するための最も望ましいのは、システム生物学とネットワーク機能のレベルであると信じています。神経回路がどのように動作し、どのような機能を実行するかを理解することは、欠陥のある脳領域を修復または回避するのに役立つ重要なリハビリテーション戦略を含む、診断および治療戦略を考案するのに役立ちます。
多くの生物学的ネットワークは、小さな世界とスケールフリーの組織を持っています。現在の証拠は、脳には効率的な情報処理の基礎となる小さな世界の接続性があることを示しています。また、高度に接続されたハブとモジュール性も備えているため、スケールフリーの特性も得られる可能性があります。このような組織は、これらのネットワークがどのように機能するか、また機能不全がネットワーク内でどのように攻撃し伝播するかを理解するためのフレームワークを提供します。
脳を計算装置として理解することは、精神医学の診断と治療に影響を与えます。重要な考慮事項は、脳がどのようにしてエラーを生成するのか、そして健康な脳が持続的な機能不全に陥ることなくそれらの間違いをどのように修正するのかということです。
推奨される読み物
バラバシ、A.-L. (2002年)。リンク: ネットワークの新しい科学。マサチューセッツ州ケンブリッジ: Perseus Publishing。
カンデル、E.R. (2006)。記憶を求めて: 新しい心の科学の出現。ニューヨーク: WW Norton & Company。
ルドゥー、J. (2002)。シナプス自己: 私たちの脳がどのようにして私たちになるのか。ニューヨーク:バイキング・プレス。モンタギュー、R. (2006)。なぜこの本を選んだのでしょうか?私たちがどのように意思決定を行うか。ニューヨーク:ダットン・プレス。 J.D. パワー、D.A. フェア、B.L. シュラッガー、S.E. ピーターセン (2010)。人間の機能的な脳ネットワークの発達。ニューロン、67、735-748。
その他の参考文献
Andrews-Hanna, J. R.、Reidler, J. S.、Sepulcre, J.、Poulin, R.、および Buckner, R. L. (2010)。脳のデフォルトネットワークの機能的解剖学的な分割。ニューロン、65、550-562。
Attwell, D.、および Laughlin, S.B. (2001)。脳の灰白質における信号伝達のためのエネルギー予算。脳血流と代謝のジャーナル、21、1133-1145。
バックナー、R.L. (2010)。予測と想像力における海馬の役割。心理学の年次レビュー、61、27-48。
Buldyrev、S.V.、Parshani、R.、Paul、G.、Stanley、H.E.、Havlin、S. (2010)。相互依存したネットワークにおける障害の壊滅的な連鎖。自然、464、1025-1028。
De Bruijn, E.R.A.、de Lange, F.P.、von Cramon, D.Y.、Ullsperger, M. (2009)。エラーにやりがいがあるとき。神経科学ジャーナル、29、12183-12186。
デコ、G.、ジルサ、V.K.、マッキントッシュ、A.R. (2011)。脳内の安静状態の活動を動的に組織化するための新たな概念。 Nature Reviews Neuroscience、12、43-56。
ヒル、B. (2001)。励起可能な膜のイオンチャネル (第 3 版)。マサチューセッツ州サンダーランド: シナウアー
株式会社アソシエイツ
Esser, S.K.、Hill, S.、Tononi, G. (2009)。徐波睡眠中の効果的な接続の破壊: 大規模モデリングを使用して、皮質ゲートの根底にあるメカニズムを調査します。神経生理学ジャーナル、102、2096-2111。
フォックス医学博士、ライクル医学博士 (2007)。機能的磁気共鳴画像法で観察される脳活動の自発的変動。 Nature Reviews Neuroscience、8、700-711。
フリストン、K.J. (2009)。機能的神経画像化におけるモダリティ、モード、およびモデル。科学、326、
399-403。
Horovitz, S.G.、Braun, A.R.、Carr, W.S.、Picchioni, D.、Balkin, T.J.、Fukunaga, M.、他。 (2009年)。深い睡眠中の脳のデフォルトモードネットワークの切り離し。米国科学アカデミー紀要(米国)、106、11376-11381。
北野博史(2002)。システム生物学: 簡単な概要。科学、295、1662-1664。
Lewis, C.M.、Baldassarre, A.、Committeri, G.、Romani, G.L.、および Corbetta, M. (2009)。学習は、休息している人間の脳の自発的な活動を彫刻します。米国科学アカデミー紀要(米国)、106、17558-17563。
リンデン、D.J. (2007)。偶然の心: 脳の進化が私たちにどのように愛、記憶、夢、そして神を与えてきたのか。マサチューセッツ州ケンブリッジ:ベルナッププレス。
メスラム、M. (2009)。計算された接続性の大胆な新しい世界で神経認知ネットワークを定義します。ニューロン、62、1-3。
レイクル、M.E. (2008)。人間の脳のマッピングに関する簡単な歴史。神経科学の動向、32、
118-126。
レイクル、M.E. (2009)。脳機能イメージングにおけるパラダイムシフト。神経科学ジャーナル、29、12729-12734。
Szalay, M.S.、Kovacs, I.A.、Korcsmaros, T.、Bode, C.、および Csermely, P. (2007)。ストレスによる細胞ネットワークの再構成:防御と薬剤設計への影響。 FEBS レターズ、581、3675-3680。
トンプソン、R. H.、スワンソン、L. W. (2010)。仮説に基づく構造的接続性分析は、脳アーキテクチャの階層モデルを介したネットワークをサポートします。米国科学アカデミー紀要(米国)、107、15235-15239。
ワッツ、D.J. (2003)。 6 度: つながる時代の科学。ニューヨーク: WW Norton & Company。 Woolsey, T.A.、Hannaway, J.、Gado, M.H. (2008)。脳アトラス – 人間の中枢神経系の視覚的ガイド (第 3 版)。ニュージャージー州ホーボーケン:ジョン・ワイリー&サンズ。
Zhang、D.、Raichle、M.E. (2010)。病気と脳の暗黒エネルギー。 Nature Reviews Neurology、6、15-28。
Zhang、Q.、Haydon、PG (2005)。神経系における膠細胞伝達の役割。神経伝達ジャーナル、112、121-125。
Zorumski、C.F.、Isenberg、K.E.、Mennerick、S. (2009)。細胞およびシナプスの電気生理学。 B. J. Sadock、V. A. Sadock、P. Ruiz (編)、カプランとサドックの精神医学の包括的な教科書 (第 9 版、129-147 ページ)。メリーランド州ボルチモア: リッピンコット・ウィリアムズとウィルキンス。
