
5 精神障害と脳ネットワーク
精神医学的症候群がどのように発生するのか、またそれらの最善の治療法を理解するには、脳がどのように情報を処理するのか、また脳ネットワークの機能不全がどのように症状を引き起こすのかを理解する必要があります。精神疾患には、1 つまたはいくつかの特定の内因性接続ネットワーク (ICN) における一次的な機能障害が関与している可能性があります。この機能不全は、他のICNに代償性の変化をもたらします。これらの変化の一部は機能を改善する可能性があり、一部は一次的欠陥を悪化させる可能性があります。重要なのは、時間の経過とともに、すべての主要な精神疾患には心の 3 つの領域にわたる機能不全が関与していると私たちが主張することです。したがって、特に本格的な病気の場合、認知、感情、および動機の欠陥を反映する症状が一般的であり、例外ではありません。このクロスモダリティ機能不全は、脳がどのように組織されているか、ICN がどのように相互作用して思考や行動を生成するか、そして神経系全体にわたるシナプス相互作用と脳の可塑性がどのように個人の利益と不利益の両方に作用するかを反映しています。これらの概念は、精神科医が精神障害の性質をより厳密に理解するのに役立つと私たちは信じています。さらに、特定の脳システムの機能不全がどのようにして特定の症状を引き起こすのかを理解することは、最終的には、神経系を共有する症状に基づいて同一個人における多数の併存疾患の過剰診断を含む、精神科診断における現在の問題のいくつかを単純化し、修正するのに役立つと信じています。心を構成するネットワーク。たとえば、どの ICNS が主に機能不全に陥っているのか、またどの ICN が欠落または不適切な入力の補償に関与しているのかを理解することで、精神病性障害についてより深く理解できるようになる可能性があります。この章では、脳ネットワークと精神疾患との関係の概要を説明します。精神医学的症候群を脳システムに関連づけるこのアプローチはまだ初期段階にあるため、ここで議論されている概念は絶対的な事実を述べたものではなく、むしろ現場に浸透している考え方と私たち自身の解釈と推測を表しています。
精神障害と精神的誤りの修正の欠陥
この議論を始めるにあたり、精神疾患は最終的には神経の恒常性と、脳が生成するエラーをどのように修正するかの問題を反映している可能性があることを再度強調します。この問題については本書の前半で言及しましたが、ここでこのアイデアをさらに詳しく検討します。人間の脳は、外界と内界に関する膨大な量の情報を処理し、追跡する複雑なコンピューティング デバイスです。これを効率的に行うには、脳はショートカットを使用してエネルギーを節約し、処理時間を短縮する必要があります。無意識の感情や動機は脳のショートカットの例です。脳は高次の思考においてもショートカットを使用します。たとえば、私たちは日常的に限られた情報に基づいて、あいまいな状況について推論や結論を導き出します。これらの結論は、いったん導き出されると、たとえ矛盾する証拠に直面したとしても覆すのが難しい場合があります。私たちの「認識」は、ほとんどの場合、新しい感覚入力と過去の経験の組み合わせを反映しています。脳は古いものと新しいものを接続するショートカットを使用するため、結果として得られる計算は必ずしも事実に基づくものではなく、中立的な観察者が経験する現実に基づくものでもありません。これは、脳が「間違い」を犯しやすいという事実に寄与しています。その間違いは、ほとんどの場合軽微なもの(例:単純な誤解、誤った記憶、誤解)ですが、場合によっては深刻なものになる可能性があります(大きな感情的な過剰反応、精神異常な考えなど)。 。
脳への損傷は、欠陥のあるICNSが情報を効率的に処理できなかったり、欠陥を補うか情報が欠陥であることに気づかずに正常に機能しているICNに誤った情報を送信したりする結果として、エラーの発生を拡大させます。私たちの脳は最終的にすべての「現実」を作り出しますが、場合によっては、現実として認識されているものが、他の人の観点からは単に間違っていることもあります。重要なのは、健康な脳はエラーを修正する能力に優れているということです。誤り訂正には、新しい入力と結果に基づいて情報処理を更新することが含まれ、通常は脳の可塑性と、反復的な強化ベースの学習形式の利用など、新しい情報を学習して記憶する能力が関係します。私たちの脳は、受け取る情報と、受け取ると期待される情報 (つまり、現実) を常に比較します。意見の不一致がある場合、脳はその不一致の大きさを修正しようとします。脳の可塑性は、内部および外部の現実を更新するための基盤を提供します。更新により、常にではありませんが、通常は 2 つの世界の一致がもたらされます。
私たちは、人間は、精神障害のない人も含めて、世界について奇妙な考えを抱いたり、機嫌が悪くなり、異常な動機を経験することがよくあると主張します。我々は以前、成人の約4分の1が、魔術的な考え(迷信や特異な信念など)、数を数えたり番号を付ける儀式、買いだめ行動などを含む重大な強迫観念や強迫行為を報告していることを指摘した。成人の約 3 分の 1 が、何らかの迫害的な考え方、たとえば、自分は他人から意図的に不当な扱いを受けている、他人によって人生を抑圧されている、他人から嘲笑の対象になっているという信念を報告しています。これらの人々のほとんどは精神疾患を患っていないか、少なくとも強迫性障害(OCD)または精神病性疾患を患っているとして臨床上の注目を集めていません。彼らは、自分たちの独特の思考や行動を、生活の中で機能するのに十分な制御下に保つことができます。理由は不明ですが、行動が精神障害の閾値に達している人は、思考、気分、モチベーションの誤りを修正したり、調整したりすることができません。これらの間違い、特に感情的なものは時間の経過とともに悪化し、本人や周囲の人々にとって不快になる持続的で異常な行動につながります。
認知神経科学者のポール・フレッチャーとクリス・フリスは、精神医学の観点から誤り訂正の問題について議論しました。彼らは、関連する入力と無関係な入力を区別できないことが、精神障害を持つ p にとって大きな問題であると仮定しています。したがって、病気を患っている人は無関係な情報に基づいて行動する傾向があり、自己監視に欠陥があるように見えます。たとえば、統合失調症では、問題は論理的推論それ自体の欠陥ではなく、むしろ確率論的推論の欠陥、つまり、信念を支持する弱い情報を過大評価する一方で、信念に反する強力な証拠を軽視する傾向であるように見えます。統合失調症の人は、誤って重み付けされた情報を扱うことで、自分自身や世界について不正確な結論に達し、新しい入力に基づいて自分の信念を更新することができなくなります。これは一種の学習または可塑性の欠陥です。興味深いことに、統合失調症にはドーパミン伝達系の欠陥が関係していると考えられることが多く、ドーパミンの主な機能の 1 つはエラー予測信号を生成することです。したがって、統合失調症患者の誤り訂正の問題は、ドーパミンを介した伝達を使用する神経回路に問題がある可能性があります。これらの欠陥は、作業記憶や明示的学習に影響を及ぼし、使用の困難につながる可能性があります。
修正手段としての新しい情報。
他の精神疾患を持つ人も誤り訂正能力に欠陥を示しますが、その欠陥の特徴は統合失調症に見られるものとは異なるようです。 Farshad Mansouriらは、認知機能検査の研究に基づいて、大うつ病患者は誤り発見と誤り訂正ができると報告している。ただし、精神的なエラーを処理して修正するには、より多くの時間と神経細胞の努力が必要です。この時間的遅延の間に、まだ修正されていないエラーが新しい学習に組み込まれるまでの時間が長くなり、欠陥が永続化して悪化します。問題は、うつ病の人が常に情報を不正確に評価するということではありません。むしろ、修正システムの動作が遅すぎて、間違いが思考や認識に根付いてしまいます。うつ病の人は、ネガティブな感情的バイアスのために状況を不正確に評価することも知られており、これがエラーの生成と修正の問題を複雑にします。ネガティブに帯電した出来事がポジティブな経験よりも優先され、これも確率論的推論における感情に基づくバイアスを反映しています。
OCD は、エラー処理で発生する問題の 3 番目の例を示しています。この障害を持つ人は、関連する情報を正確に評価でき、その情報を十分に速く処理できます。しかし、彼らは、無関係な入力が関連するアイデアを妨げるのを防ぐのが難しいようです。これは一種の注意力と認知力の柔軟性の問題です。実際、関連性のある思考と無関係な思考や認識が絡み合い、高い感情的要素(不安や不快感)をもたらします。 OCDで観察される問題は、ある意味、統合失調症で観察される誤り訂正の問題に似ているように思えます。これには、物事について魔法のような信念を抱いていることが含まれます(たとえば、悪い結果をもたらす自分自身の思考や行動の力)。これは、妄想的な考えと重度の強迫観念的な考えを区別するのが難しい場合があることを示す臨床観察と一致しています。
脳が精神的なエラーを修正できない場合、さまざまな症状が発生する可能性があります。統合失調症の人は、情報の関連性を正確に判断できないために、被害妄想を発症することがあります。うつ病患者は、自分の価値について誤った認識を持っている可能性があり、その考えを適時に修正できないと、その考えが思考に根付き、罪悪感や死の思考につながります。 OCDを持つ人は、無関係な入力がエラー修正を妨げる可能性があり、関連する情報に戻ろうとし続け、その過程で習慣的な強迫観念や強迫行為を発症する可能性があります。時間が経つにつれて、誤った考えは日常的な思考に組み込まれ、より執拗で習慣的なものになります。この状況は心理社会的ストレスによってさらに悪化します。ストレスが増大すると、人間や他の種はエネルギーを節約するために脳が使用する方法としての反応がより習慣的(暗黙的)になります。
精神障害を持つ人はなぜ精神的な間違いを正すことができないのでしょうか?
誤り訂正と精神疾患におけるその役割は複雑なテーマであり、まだ詳細には理解されていません。私たちは、エラー修正は脳が効率的に機能するために使用する一連の重要なメカニズムを反映しており、これらのメカニズムが正しく機能しない場合に問題が発生すると考えています。このセクションでは、誤ったエラー修正に寄与する可能性のある要因について推測します。これらは表 5-1 で強調表示されています。
エラー修正には、多くの場合、感情、動機、知覚、つまり神経処理のより原始的なモードにおけるエラーをオーバーライドする高次の脳システムの能力が必要です。したがって、誤り訂正とホメオスタシスの欠陥の原因の 1 つは、トップダウン制御の失敗である可能性があります。これは複数の方法で発生する可能性があります。たとえば、知的発達が障害されると、定期的に浴びせられる複雑な情報を処理する脳の能力が制限される可能性があります。知的発達の障害に伴う認知の柔軟性の低下と認知制御の欠陥は、一般知能(標準的な IQ テストで測定)が低い人は、統合失調症、薬物乱用、大うつ病などのいくつかの主要な精神疾患のリスクが高いという観察に寄与している可能性があります。同様に、幼少期の重度のストレス(栄養失調、虐待、育児放棄など)から生じる発達上の問題は、神経損傷や異常な学習の結果、高次皮質機能を妨げる可能性があります。皮質主導のトップダウン制御の問題は、人生の他の時期にも、認知、注意、感情に関与するICN間の機能不全や切断を引き起こす特定の脳病変(外傷性脳損傷や脳卒中など)や神経変性疾患から生じる可能性がある。 、そしてモチベーション。
————————
表 5-1 なぜ自己修正ができないのか?
変更されたトップダウン制御
神経発達の問題 (1Q/初期ストレス)
病変/神経変性(細胞喪失、皮質の薄化)
感情/動機/認知間の断絶 (シナプス/接続の問題)
感情、動機、または認知システムにおける主要な欠陥
誤った学習(可塑性の問題)
モチベーションの横取り(薬物/衝動/アイデア)
————————
トップダウン処理の失敗に加えて、一部の個人は、感情的および動機付けのシステムに主要な広範な問題を抱えている可能性があり、そのために高位センターが誤った情報を修正することが困難になります。たとえば、幼児期の虐待やネグレクトは過剰反応性のストレス反応と関連しており、比較的無害な出来事が不適切な感情的および身体的反応を引き起こす可能性があります。幼少期のストレスの多い出来事が、特に特定の遺伝的素因と結びついた場合、感情の発達を妨げ、結果として皮質の制御を無効にするストレスと感情システムを引き起こす可能性はあります。これは、初期のストレスを介した皮質発達の変化と組み合わされて、感情の調節に重大な困難を引き起こす可能性があります。同様の考慮事項は、モチベーションと報酬システムに影響を与える可能性があります。たとえば、特定の薬物を乱用する個人は、動機顕著性システムに変化が生じており、そのためシステムの自己修正が困難になっています。乱用された薬物はすべて、脳の動機と報酬システムに深刻な影響を与え、これらのシステムの構造と機能に長期にわたる持続的な影響をもたらし、その影響は薬物使用期間をはるかに超えて持続することがよくあります。通常、新皮質は、動機システムによって生成される衝動を制御します。薬物乱用は、これらのシステム間の機能的接続のバランスを取り直す可能性があり、皮質接続の有効性を低下させ、報酬やストレスを軽減するためのより健康的な経路の代わりに、不十分に調整された感情に基づく行動を可能にする可能性があります。この動機付けシステムの機能的および物理的な再配線は、薬物乱用症候群の治療が困難であることが多く、再発を繰り返す可能性があるという事実に寄与している可能性があります。
興味深いことに、薬物乱用は、動機を奪い、規制を困難にする唯一の方法ではありません。人間が抽象的思考の能力を進化させるにつれて、抽象的思考を動機として利用する能力も発達しました。したがって、人間は、より生理的な欲求 (食べ物、性別、生存) に加えて、抽象的な概念を使用して行動を推進することができます (たとえば、試合に勝とうと努力したり、政治的大義のために戦ったりするなど)。極端に考えられると、抽象的な信念は論理を無効にする可能性があり、政治的または宗教的な自殺で見られるように、人間は信念のために身を滅ぼすこともあります。実際のところ、私たちはおそらく、自分自身の思考にこれほど「誘惑」される唯一の種です。ここで重要なのは、特定の思考が私たちの脳の処理に深く根付き、薬物乱用と同様に、合理的な制御を無効にしてしまう可能性があるということです。
精神医学的分類とエンド表現型を再考する
以前にも述べたように、私たちは主要な精神疾患は、思考、感情、動機という心の 3 つの領域にわたる機能不全に関係していると考えています。ただし、これは、特定の病気の機能不全を引き起こす主な病理が、これらの計算システムを構成する ICN に同様に影響を与えることを意味するものではありません。たとえば、実行機能に関与する皮質中枢は、他の脳システムに異常が生じると機能不全を起こしやすくなります。アルツハイマー病では、通常、実行機能 (計画と意思決定) に問題が生じますが、初期の病理が記憶システムを攻撃し浸透します。高次機能障害は二次的な症状を表します。病気に関連して個人に悪影響を与える可能性があることを考慮することも重要です。病気が持続し進行するにつれて、1つのシステム内で発生した脳の可塑性エラーがどのように伝播するかを精神的に説明します。誤った処理はより結晶化して蔓延し、時間の経過とともに脳回路の程度が拡大する可能性があります。薬物乱用障害は、時間の経過と薬物の反復使用によって病気が進行するにつれて、拡張する神経回路が役割を果たすようになるという、この概念の有益な例を表す可能性があります。これについては、この章で後ほど詳しく説明します。今のところ、私たちは心の三要素構造に従って精神医学的分類について考えることが有用かどうか、つまり、認知、感情、または動機における一次的機能障害を反映するものとして精神疾患を概念化することが有用かどうかを考えてみたいと思います。このような分類は、これらの主要な心の構成要素のうちの 1 つに機能不全の主要な場所を定義しようとするものであり、病気の初期の症状を定義したり、病気が時間の経過とともにどのように進行して心の 3 つの要素すべてに影響を与えるかを理解するのに役立つ可能性があります。 。このようなスキームに従って病気の段階分けができれば、まだ本格的な機能不全を発症していない高リスクの個人を含む、病気の段階にターゲットを絞った合理的な治療およびリハビリテーション戦略を設計することが可能になるかもしれない。このようなスキームは、人格障害(または薬物乱用障害)を背景とした重度のうつ病の治療が、他の精神疾患を持たない個人の「原発性」うつ病よりもなぜ難しいのか、そしてなぜそれが不適切であるのかを理解するのにも役立つかもしれない。これら 2 つのシナリオで同様の治療戦略を使用します。
表 5-2 は、そのような分類スキームがどのように構築されるかを示す簡単な例を示しています。この例では、認知症疾患、精神遅滞症候群、およびせん妄が、高次レベルの処理における重大な欠陥に起因する主要な認知機能障害とみなされます。これらの障害はすべて、学習、記憶、推論に重大な問題を示します。せん妄は覚醒とより原始的な脳機能障害を強く伴う複雑な症候群であるため、このグループにせん妄を含めることについては確信が持てません。同様に、精神障害やおそらく強迫性障害も、病気の過程の初期に存在する、主要ではあるが異なる認知の欠陥が関与しているとみなされるのではないかと考えています。精神病性障害を認知症および発達性認知障害と並べることは、後者の障害が精神病性症状と頻繁に関連していることと一致するであろう。さらに、あらゆる精神病性障害にはさまざまな程度の神経心理学的機能不全が伴い、これらの欠陥は最初のエピソードの発症時に存在することが多いという証拠があります。また、そのようなスキームでは、現在どのように障害を分類しているかに関係なく(例:統合失調症、双極性障害、または精神病性大うつ病)。これはまた、精神病の存在により、特定の気分障害が非精神性気分障害よりも統合失調症とより密接に一致することを意味します。これは、精神障害全体に共通する遺伝学とより一致している可能性があり、精神病的思考の発生につながる、またはそれに関連する認知の欠陥を対象としたリハビリテーションの戦略の開発につながる可能性がある。この分類は、精神障害における認知欠陥が仕事関連障害の主な要因として重要であることとも一致するであろう。
————————
表 5-2 精神障害と心: 主要な欠陥と障害?
原発性認知障害?
作業記憶/注意/実行機能 (PFC-頭頂ネットワーク)
・認知症、精神薄弱、せん妄
・精神障害
・強迫性障害?
原発性感情障害?
ネガティブな感情(mPFC-pgACC-拡張扁桃体ネットワーク)
・気分障害
・不安障害
原発性動機障害?
VTA-側坐核-PFCネットワーク
・薬物乱用・アルコール依存症
・摂食障害
・パラフィリア
・パーソナリティ障害
————————
このアプローチの問題は、個々の患者における精神病症状の有無の判断、さらには精神病症状の構成要素の判断にある可能性があります (第 1 章)。しかし、臨床現場では、精神病性の特徴を伴う障害は通常、非精神病性疾患とは区別されるものとして見られ、扱われるため、私たちはこれを大きな問題とは考えていません。気分障害の場合、あまり明確になっていないのは、特定の患者のうつ病または躁状態のエピソードが精神病とも関連しているかどうかです。気分障害のある人の中には、精神病症状の有無にかかわらず、一定期間の病気を示す場合があり、これが提案されている概念化を複雑にしています。しかし、私たちは、ある人がいったん明白な精神病を発症すると、ほとんど原因に関係なく、その人は非精神病型の障害よりも複雑な病気を患っていると主張します。
他の障害、特に非精神性気分障害や不安障害は、拡張扁桃体、報酬系、および関連するICNの機能における遺伝的および後天的変化を伴う、感情システムの主要な欠陥を反映していると見なされる可能性があります。これと一致して、うつ病の人はポジティブな感情を維持することが非常に困難であり、これは、楽しい経験を処理する際に前頭葉皮質と側坐核を含む回路の活動を維持できないことと相関しています。脳の回路が共有されていることが、気分症候群と不安症候群の大部分の併存症の原因である可能性があります。繰り返しますが、これらの病気が進行するにつれて、認知システムや動機付けシステムに二次的に関与する可能性がありますが、主な欠陥は感情システムに見つかる可能性が高くなります。
一部の精神障害は、モチベーション経路の一次欠陥から生じると見なすこともできます。薬物乱用障害は、乱用薬物の既知の薬理学と、腹側被蓋-側坐核ネットワークを標的とする薬物の傾向に基づく最も明確な例です。また、摂食障害、パラフィリア、ギャンブル障害、そしておそらくパーソナリティ障害がそのような計画に当てはまるかどうかも推測します。なぜなら、これらの障害グループはそれぞれ、異常な動機に関する大きな問題を示すからです。
これらの線に沿って精神疾患を再分類するのは明らかに時期尚早であるが、このような概念的スキームは、脳ICNと、おそらくは疾患の遺伝学および生物学とより密接に関連する中間表現型を定義するのに役立つ可能性がある。一例として、認知症症候群の病理および臨床症状における脳 ICNS の役割に関する最近の研究に戻ります (第 2 章)。これらの症候群では、臨床表現型は特定の ICN 内の機能不全によって引き起こされますが、機能不全の根底にある分子機構は表現型間で共有される可能性があります。たとえば、アルツハイマー病の通常の臨床表現型には、デフォルト ネットワークに対する一次攻撃が関与しているように見えますが、頻度は低いですが、アルツハイマー病の分子病理学的特徴であるプラークや線維変化も、行動変異型前頭側頭型認知症の臨床症状を引き起こす可能性があります。これは、プラークともつれが別の ICN (感情顕著性システム) を攻撃するときに発生します。より多くの場合、行動変異型前頭側頭型認知症に伴う神経変性には、アルツハイマー病で見られるものとは異なる分子病理が関与しています。特定の個人において特定の ICN が脆弱になる原因はまだ明らかではありませんが、デフォルトのネットワーク処理に必要な高い活動とエネルギーがアルツハイマー病において重要であると考えられ、これは将来の治療と予防を考案する際に役立つ可能性がある 1 つの手がかりを提供します。戦略。
これらの概念を精神疾患に拡張すると、Jaak Panksepp が提案したように、動物や人間で観察される一次感情に基づく内部表現型を探すこともできます。私たちが暫定的に一次性感情障害(非精神性気分障害および不安障害)と呼んでいるものは、主に強い否定的な感情バイアスを反映しています。うつ病の場合は悲しみ/敗北、不安障害の場合は恐怖/不安、そしておそらく怒り/他の障害で激怒する。躁状態の高揚感は例外とみなされるかもしれませんが、ここでも高揚感が過敏症や怒り、躁状態と鬱状態の混合状態に堕落することがよくあります。特定の気分状態につながる神経計算は理解が進んでおり、実際の結果または知覚された結果と内なる期待との比較が含まれているようです (表 5-3)。結果が期待を下回ると、脳システムがリセットしようとして感情が引き起こされます。経験される特定の感情は、個人の感情的ICNの内部バイアスを反映している可能性があり、敗北/損失の認識は悲しみを引き起こし、脅威の認識は不安を引き起こし(逃避)、不公平の認識は激怒を引き起こします(闘争)。感情的なトリガー自体は、認識された危害、失敗、拒絶、曖昧さなど、いくつかのテーマを中心に展開しているようです。曖昧さ(不確実性)は、最も具体性が低いため、人間にとって最も対処が難しいものと考えられます。曖昧さは、次のことを必要とする感情的な引き金になることもあります。
————————
表 5-3 否定的な感情的バイアス

————————
分析にはより多くの脳の労力が必要となるため、実行システム、感情システム、モチベーションシステムの間の高度な調整が必要になります。
この議論は、私たちの脳の「期待」を決定する要因について重要な疑問を投げかけます。これらの概念については第 4 章で説明しましたが、ここではテーマを少し拡張します。最も原始的なレベルでは、人間は生存と生殖に影響を与える衝動(食べ物、性別、保護など)によって動機付けられています。人間はまた、生存や進化には必要ではないが、魅力的で有利なものである欲求によって動機付けられています。このような場合、脳の微積分は、知覚された結果や行動がコストに見合うかどうかの判断を反映します。この計算では、社会的承認などの概念や、魅力的だと思うものについての私たち自身の抽象的な概念さえも考慮に入れることができます。私たちの性格に固有の特徴、特に C. ロバート・クロニンジャーらが気質と呼んでいる性格の次元が、これらの決定における主要な変数であるようです。クローニンジャーは、気質を、私たちの感覚によって知覚される比較的単純な刺激によって引き起こされる習慣やスキルと定義しています。気質は、危害回避、新奇性追求、報酬依存、持続性などの特徴の観点から説明されており、これらは遺伝性があり、発達的に安定しており、主に社会的学習によって引き起こされるものではありません。これらの特性は、私たちの注意を引くものや、感情的な顕著性が高いものを判断するのに役立ちます。興味深いことに、第 4 章で説明したように、神経画像研究は、これらの気質特性が、動機付けの ICN の一部である特定の神経回路の活動を反映していることを示しています。
ICNS と精神障害について私たちは何を知っていますか?
認知障害: 精神病
ICN が主要な精神疾患にどのように関与しているかはまだ理解されていないと言っても過言ではありませんが、特定の脳病変が精神症状を引き起こす行動神経学から学んだ教訓は、方向性を示すのに役立つ可能性があります (第 1 章を参照)。したがって、精神病は私たちの認知システムの構成要素に関係する主要な欠陥として調べることができると私たちは信じています。たとえば、一部の精神病症状には、環境内の人々の妄想的な誤認が含まれます。カプグラ症候群では、自分の近くにいる人が正確な影武者に置き換わったという思い込みが生じます。特定の脳病変も妄想性誤認と関連しています。これらの病変は通常、非優位半球または両方の前頭葉に影響を及ぼし、自己を監視し、不正確な認識を修正する能力に欠陥をもたらします。臨床例には、自分がいる場所とまったく同じ複製である場所にいるという思い込みが生じる重複性記憶喪失症候群が含まれます。第 1 章で述べたように、同様の脳病変は病態失認にも関連する可能性があります。たとえば、人が手足を自分の体の一部として認識できないなどです。このような場合、右半球の損傷により原発性の神経学的欠陥(左側半身麻痺など)が生じますが、損傷を受けていない左半球が過剰に活動し、異常な説明が得られます。これらの症候群では、病変領域からの誤った入力が左半球からの過剰な一般化と結合して症状を引き起こします。カプグラ症候群の根底にあるメカニズムはそれほど確実ではないが、認知処理の欠陥が感情の変化と相互作用して症状を引き起こす可能性がある。 V.S.ラマチャンドランは、カプグラ症候群には知覚(家族の見たり聞いたり)と感情(ポジティブバランスの欠如)の間の接続の変化が関与している可能性があり、期待される感情を経験できないことが妄想的な説明につながる可能性があると示唆しました。認識された愛する人がほとんどまたはまったく感情を引き起こさない場合(または否定的な感情さえも)、左半球は、その人は愛する人ではあり得ず、代わりに詐欺師に違いないという説明を生成します。この場合、この症候群の主な原因は感情システムと認知システムの間の断絶であるが、認知システムは欠陥のある入力を理解しようとして、愛する人が自分に取って代わられたという誤った信念を育む。正確な二重。前述したように、確率的推論の大きな欠陥が妄想の中心にある可能性があります。おそらくこれに関連して、統合失調症の人は新しい情報と古い情報を区別することが困難であり、右半球の処理の欠陥が認識記憶の問題の一因となっているという証拠もいくつかあります。そして、左半球は、誤って認識された情報に対して妄想的な結論を提供しているように見えます。他の証拠は、認識記憶の低下と関連した脅威反応の障害が精神病症状の悪化に寄与していることを示唆している。
多くの研究が、精神疾患のある人およびない人における迫害的な考えに寄与する精神的要因を調査しました。人間集団全体で、迫害的な考えと猜疑心が妄想の最も一般的な形態を表しており、これらのタイプの考えは、明らかな精神疾患を持たない人々の間では比較的一般的ですが、後者の人々では、それらは彼らほど揺るぎないものや奇妙なものにはなりません。本当の妄想を持っている人の中にいます。迫害的な観念化に寄与する要因には、持続的に欠陥のある思考につながる感情的および認知的処理の問題の組み合わせが含まれるようです。迫害的な考えや妄想を持つ人は、通常、実行機能や意思決定の欠陥(例:結論に飛びつき、物事を誤解する傾向など)とともに、否定的な感情的バイアス(悲観主義、不快感、自尊心の低さなど)を示します。他人の精神状態)。問題解決や確率的推論の問題も寄与しており、奇妙な「アイデア」から「妄想」への移行の主要な要因となる可能性があります。実行機能の欠陥と結びついた否定的な感情的バイアスの影響により、注意の集中方法、状況の評価方法、帰属の方法に関する認知バイアスが生じます。重要な点は、妄想的思考には認知的根拠があるが、単なる思考の問題ではないということです。妄想的思考は認知的欠陥と感情的偏見の組み合わせを反映しています。複数の ICNS が症状の一因となりますが、認知的誤解が症状の現れ方に主導的な役割を果たし、おそらく異常な考えがどの程度定着するかを決定することになります。
精神病における複数のICNSの関与は、統合失調症などの病気を考慮するとさらに強調されます。この精神病性障害は、陽性症状(妄想、幻覚、思考障害)、陰性症状(感情鈍麻、非社交性、失語、無力症、無気力[いわゆる「5A」])、認知欠陥(選択的問題)によって特徴付けられます。とりわけ注意力、認識記憶、作業記憶など)、そして時には運動症状も伴います。興味深いことに、認知欠陥は多くの人にとって長年の問題であるようです。それらは病気の発症時に存在することが多く、時間が経っても比較的安定した状態を保ちます。それらはまた、病気の長期経過と障害のレベルを決定する上で重要な役割を果たしており、統合失調症患者が学校を卒業したり、有利な職に就くことが困難になる主な理由となっている(クレペリン早発性認知症の「認知症」)。認知障害も、迫害的な考えなど、病気に関連した異常な思考の一因となっている可能性があります。
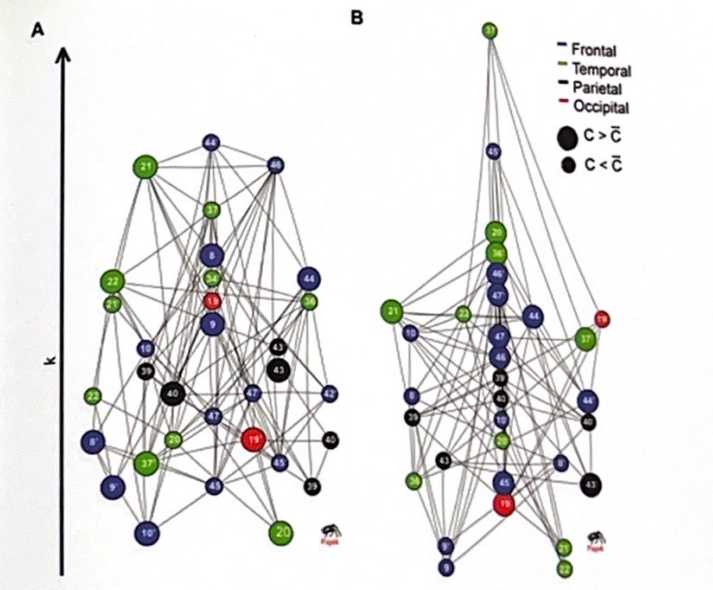
統合失調症患者の脳では何が問題を起こし、認知機能障害や症状を引き起こすのでしょうか?これはまだ進行中ですが、脳ネットワークの研究からいくつかの洞察が得られています。統合失調症患者は、海馬および新皮質の複数の部分(PFC を含む)の構造変化(細胞の縮小と損失)を伴う脳領域全体の欠陥を示します。我々は以前、脳ネットワークには、ネットワーク内の領域間の高度な局所クラスタリング(短い経路長)と、少数の長距離接続を介したより離れた領域間の効率的な情報転送を反映するスモールワールド接続性があることに注目しました。 Danielle Bassettらは、機能画像を使用して、脳ネットワークの接続性に関して健康な対照者と統合失調症患者がどのように比較されるかを調べた。統合失調症の人は、新皮質の階層構造の低下と前頭皮質ハブの接続性の低下を伴う、異常な多峰性ネットワーク組織を示します。対照的に、非前頭皮質領域は、対照では見られない高度に接続されたハブを示した。その結果、脳の構造はマルチモーダルなデータの処理効率が低下し、前頭葉に関わる一種の接続不全と一致します。図 5-1 はこの研究の概要を示し、対照と統合失調症患者における皮質ネットワークの接続性と階層の違いを強調しています。
皮質領域間のスモールワールドの接続性の変化は、情報処理に実際的な影響を及ぼします。 Van den Heuvelらは、皮質ネットワークの静止状態の経路長が知能のIQ測定値と逆相関することを発見した。つまり、静止状態の経路長が長いほど、処理効率の低下と一般知能の低下に関連している。したがって、統合失調症における階層的処理の切断と喪失、特に右側の前頭頭頂ネットワークが関与することは、作業記憶、実行機能、新情報と古い情報の区別において観察される困難の原因となっている可能性があり、後者はおそらく、情報に関する欠陥のある前提を修正する際の問題に寄与している可能性がある。自分自身と外の世界。
————————図 5-1
————————
図 5-1 連結性マップ: 対照と統合失調症患者。この図は、機能的 MRI データに基づいて、対照被験者 (A) と統合失調症患者 (B) の間の皮質結合性の違いを示しています。この図は、統合失調症患者の前頭葉における局所的なクラスタリングの変化と、皮質領域を結ぶ長い経路長を示しています。 (Bassett et al.、2008 の許可を得て転載。)
————————
デビッド・ルイスとロバート・スウィートは、神経回路の観点から統合失調症について説得力を持って議論し、作業記憶と聴覚処理の欠陥を強調しました。作業記憶の欠陥には、集中して思考を維持する能力に重要な領域である背外側前頭前野 (dIPFC) の異常が関与しています。作業記憶の負荷が低いタスクを行うように依頼された場合、統合失調症の被験者はそのタスクを実行できますが、対照と比較してdIPFCの活性化レベルが高くなります。これは、より高い認知要求と、タスクを実行するためにより多くのエネルギーを使用する必要性を反映しています。作業記憶の要求が高い場合、統合失調症の人は dIPFC 活性の低下を示し、課題遂行に困難を経験します。これらの機能的欠陥に加えて、統合失調症患者は、dIPFC の錐体ニューロンおよび特定の種類の GABA 作動性介在ニューロンに構造異常を持っています。後者の細胞は、カルシウム結合タンパク質であるパルブアルブミンを発現する高速スパイク抑制性ニューロンの一種です。
これらの介在ニューロンは、錐体ニューロンの発火の調節を助ける上で重要な役割を果たし、脳領域内および脳領域間の機能的接続をサポートする脳リズムの同期を助けます。特に、統合失調症の人は、これらの介在ニューロンによって調節されるベータ(1秒あたり15〜30サイクルの活動)および(1秒あたり30〜80サイクルの活動)周波数範囲の作業関連リズムの低下を示します。これらのタイプの電気リズムは、主要な興奮性 (錐体状) ニューロンの出力を組織化し、脳領域内および脳領域全体にわたるタスクの実行に関与する細胞集合体を選択するのに役立ちます。統合失調症患者の活動性の低下は、皮質領域間、特に前頭葉領域と頭頂葉領域の間に接続不全(経路長の延長)があり、この疾患における認知症状の一因となっているという考えと一致している。これはまだ研究中ですが、統合失調症の一側面には dIPFC 処理の異常が関係しているようです。
重要なのは、統合失調症における皮質欠陥はdIPFCに限定されず、ガンマリズムの変化が新皮質全体で見られることです。たとえば、ルイスとスウィートは、他者の表現された(話された)感情を含む聴覚入力の解釈に問題を引き起こす、支配的な側頭葉のヘシュル回の変化についても説明しています。この領域および言語 ICN に関与する他の領域の欠陥は、思考障害にも寄与する可能性があります (例: 接線性、脱線)。同様に、統合失調症の人は、認識記憶課題を実行するときに、特に非優位半球における前頭前皮質と頭頂葉皮質の活性化が減弱しています。広範な皮質欠陥の存在は、統合失調症で観察される集中力の欠陥の主な原因である可能性があります。最近の大規模な神経画像データのメタ分析では、これらの考えの裏付けが加えられ、統合失調症患者は、認知制御タスクを行うときに、対照と同じ皮質および皮質下のネットワークを活性化するが、維持に必要な領域の活性化が低下している点で異なることが示されています。コンテキストとタスク、認知制御問題の一種。
他の研究では、統合失調症の人はデフォルトシステムが過剰に活動しており、注意を集中するよう求められたときにデフォルトモードから移行するのに問題があることがわかっています。集中力と作業記憶の欠陥は、統合失調症に関連する内的世界と外的世界の曖昧さの主な原因である可能性があり、より高次の認知的欠陥や否定的な感情的バイアスと組み合わさると、妄想や幻覚を引き起こす可能性があります。これと一致して、精神病(統合失調症と双極性障害)の遺伝的危険因子を調べた最近の研究では、リスクが左右のdIPFC間の接続性の低下(おそらく作業記憶の問題を反映している)と海馬(脳の一部)間の接続性の強化に関連していることが判明した。デフォルトのシステム) と dIPFC。正常な被験者は、海馬とdIPFCの間の直接的な接続をほとんど示さなかった。興味深いことに、遺伝的リスクは扁桃体、海馬、PFCの間のより強い接続性とも関連しており、これはおそらく精神症状に関連する感情的な偏りを反映していると考えられます。したがって、認知処理に関与する皮質系の一次欠陥が、統合失調症の陰性症状と陽性症状の両方を引き起こす可能性があります。これらの認知欠陥の一部は、認知システム、感情システム、動機システムの間で不均衡なコミュニケーションを引き起こす可能性があります。最近の研究では、抗精神病薬による治療がデフォルトの ICN 内の活動を調節するのに役立つ可能性があり、腹側線条体と海馬の血流の初期変化 (治療の最初の週) が良好な臨床反応を予測し、前側線条体の変化を伴うことが示唆されています。帯状皮質と前頭皮質はプロセスの後半(6週間の治療後)に発生します。これは、心の根底にある神経回路の相互依存性を再び浮き彫りにします。
私たちの考え方では、「認知」障害の中に精神病性気分障害と統合失調感情障害の複雑症候群を含めます。これは、これらの障害に関連する認知障害と、より攻撃的な経過と一致しています。しかし、これらの疾患では神経回路の変化があまり明確に定義されていないことを我々は認めています。 Jonathan Savitz と Wayne Drevets は、2009 年の包括的なレビューで、双極性障害の被験者は、海馬、眼窩 PFC、腹側 PFC を含むいくつかの脳領域の体積が減少し、背側 PFC の代謝低下が見られると結論付けました。研究者らは、双極性障害の被験者は、内側PFC、扁桃体、海馬、腹側線条体、視床下部、脳幹などの「内臓運動感情調節ネットワーク」に関与する大うつ病性障害の被験者といくつかの異常を共有していることを強調した。この回路については、この章で後ほど詳しく説明します。
強迫性障害: 認知障害ですか、それとも感情障害ですか?
OCD は通常、不安障害の 1 つであると考えられており、この病気には感情的な要素が強いことが強調されています。不安が重要な症状であることは明らかですが、OCD を他の不安障害とは分けて検討し、この障害の認知的側面がどのように生じるのか、また認知症状と感情的症状の関係に焦点を当てます。 OCD は、個人が反復的で侵入的な思考 (強迫観念) を持ち、多くの場合、関連する否定的な感情を和らげるために実行される反復的な運動行動 (強迫行為) を伴う不均一な状態です。大規模なメタ分析では、OCD の症状は 4 つの主要なテーマに集中していることがわかりました。それは、対称性 (儀式の繰り返し、順序付け、数の数え方)、禁じられた考え (通常、性的、宗教的、または攻撃的な内容を含む)、清潔さ (汚染への懸念) です。 )、買いだめ行為。 OCD に最も特徴的なものですが、これらの症状は他の精神疾患でも観察され、明確な精神疾患を持たない人にも見られます。しかし、非臨床集団であっても、これらの症状は苦痛を引き起こす可能性があります。多くの場合、否定的な感情 (不安や気分の落ち込み) がそのような症状に関連しており、臨床ケアを求める原動力となります。
OCDに関連する脳の変化には、線条体(大脳基底核)とPFC、特に感情の入力が受信、処理、制御される腹側および眼窩PFCを接続する経路が関与しているようです。これらの回路は、皮質が行動に対して抑制制御を発揮し、より柔軟な行動パターンを駆動できるようにするため、重要です。 OCD は、この線条体皮質回路の構造的および機能的変化、特に眼窩前頭皮質に関わる変化に関連しています。
この領域は、否定的なフィードバックを使用して非効果的または非生産的な行動を変更する学習形式である逆転学習にとって重要です。これらの観察は、OCD の認知的側面に注目を集め、特定の形式の学習における欠陥が臨床表現型の主な原因である可能性を高めています。興味深いことに、OCD患者とその臨床的に影響を受けていない近親者は、反転学習中に外側眼窩前頭皮質およびその他の皮質領域の活性化の低下を示し、反転学習の欠陥がこの障害の内部表現型である可能性を示唆しています。反転学習の欠陥がどのようにして生じるのかは明らかではありませんが、調節性モノアミン神経伝達物質の変化が関与している可能性があります。 OCD における主要な認知欠陥には、少なくとも、逆転学習の問題、思考の間で注意を移すことの困難、および全体的な思考の柔軟性のなさが含まれるようです。意思決定や計画の問題など、実行機能の欠陥もこの障害に関連しています。
さまざまな OCD 症状が、前部線条体視床回路内の接続の増加と明確な神経相関を持っているという証拠がいくつかあります。例えば、症状を誘発する課題を実行するとき、浄化の儀式を行っている人は腹内側 PFC と右尾状核の活性化が増加していますが、溜め込み行動をしている人は左の中心前回と右の眼窩前頭皮質の活性化が増加しています。チェック儀式を行っている人は、被殻/淡蒼球、視床、および新皮質の背側面の間の接続性の増加を示します。興味深いことに、OCD患者は外側PFCと背側線条体の間、および腹側線条体と腹側被蓋野の間の機能的接続性の低下も示しています。後者の発見は、OCD患者にとって意識的な認知制御が困難であり、誤り訂正が欠陥がある可能性がある理由を示唆しています。
OCD に関する発見は、臨床的機能不全につながる認知システムと感情システムの間の重要な相互作用を再び強調しています。さらに、OCDにおける背側線条体と腹側線条体の両方の関与は、異常な思考を運動出力、つまり強迫行為や儀式の形で現れる暗黙の習慣主導の行動に変換できる回路を提供します。これに基づいて、OCD は認知処理に重大な問題を抱えており、したがって一次性認知障害のサブタイプとして分類するのがより適切である可能性があることを示唆します。ただし、意識への望ましくない思考の侵入は一種のフィルタリング問題を反映している可能性がありますが、OCD 患者が他の認知障害に関連する作業記憶の問題を示していることはあまり明らかではありません。
主要な精神障害としての不安障害
不安障害は、過剰かつ不適切な恐怖を伴うさまざまな病気のグループです。その他の否定的な感情症状、特に抑うつ気分がしばしば観察され、症状と治療の両方の点で、気分障害と不安障害の間にはかなりの重複があります。不安は、恐怖の一次感情と最も密接な関係がある一方で、いくつかの点で恐怖とは異なることを考慮することも重要です。恐怖は通常、認識され、より明確に定義された脅威に対する迅速な反応ですが、不安ははるかに永続的で、特定の外部の合図や状況と直接結びつかない場合があります。不安障害で観察される恐怖は、個別のもの(恐怖症)および/または他の人(社会恐怖症)に向けられる場合があります。それは、広範で比較的集中力のないもの(全般性不安症)である場合もあれば、さまざまな生活上のストレス要因と時間的に関連している場合(急性または心的外傷後ストレス)、または明確または不明確な引き金を伴う個別のエピソードとして発生する場合もあります(パニック発作)。さまざまな形態の不安障害がどのように、そしてなぜ生じるのかは不明ですが、すべての形態が脳の恐怖回路に関与しているようです。したがって、扁桃体とその分散接続が中心的な役割を果たします。
不安の神経科学は、げっ歯類における十分に特徴づけられた恐怖モデルの存在によって大きく助けられてきました。この回路については第 4 章で詳しく説明し、恐怖を処理するための 2 つの経路の存在を強調しました。1 つは刺激の迅速かつ無意識的な評価を可能にする皮質下の経路、もう 1 つは新皮質と意識的な感情を含む経路です。さらに、げっ歯類の研究では、人間の不安に似た、慢性的であまり方向性のない恐怖反応が、終端線条(BNST)の床核とBNSTの殻(外側)領域を含む、より分散した拡張扁桃体回路を必要とすることが実証されている。側坐核。より拡張した扁桃体がこのプロセスにどのように関与しているかは完全には理解されていないが、BNSTの扁桃体中心核の出力末端からのコルチコトロフィン放出因子(CRF)の放出が関与しているようである。これらの研究では、回路は明らかに共有されていますが、BNSTは恐怖と不安を区別する上で重要な役割を果たしています。また、特に文脈に関して、恐怖に基づく学習における拡張扁桃体と海馬の間の相互作用をさらに強調したいと思います。図 4-4 (第 4 章) は、恐怖と不安に関与する神経系の概要を示しています。
人間の不安障害では脳の回路に何が起こっているのでしょうか?この質問に対する完全な答えは不明ですが、不安障害のスペクトルに寄与していると考えられる情報処理にはいくつかの種類があるようです。 Daniel Pine らによって説明されているように、不安には注意を払う要素があり、それによって個人が認識された脅威に方向を向くことができるようになります。このプロセスの後には、社会的および生理学的反応を含む脅威の評価が続きます。不安障害には学習と記憶の強力な要素もあります(げっ歯類の恐怖条件付けに似ています)。多くの点で、これらの処理コンポーネントは動物で観察されるコンポーネントに似ており、類似の脳回路を反映している可能性があります。したがって、扁桃体、および程度は低いですが海馬は、初期の脅威の評価を決定し、対応を方向付ける上で主要な役割を果たしていると考えられています。げっ歯類と同様に、腹側および内側 PFC の関与の証拠があり、この関与は不安処理の評価要素、およびパフォーマンスの監視と反応の予防にとって特に重要である可能性があります。不安障害は、尾状被殻や側坐核などの線条体の回路にも影響を与えます。この線条体の関与は、運動反応や習慣的反応、葛藤の監視、間違いの修正を促進するために重要であると考えられます。この回路は不安状態では過剰に活性化しますが、社会恐怖症で観察される変化は、特定の恐怖症や OCD で観察される変化とは異なるようです。
この恐怖/不安ネットワークの一部は、特定の不安障害でも構造変化を示します。たとえば、心的外傷後ストレス障害 (PTSD) では海馬、そしておそらく扁桃体が収縮するという証拠があります。海馬の構造変化により、PTSD と他の不安障害が区別される可能性があります。精神障害におけるストレスの役割については後続の章で扱いますが、ここでは、遭遇する症状の種類(例:感情の麻痺、出来事の侵入的再体験)を考慮して、PTSDを他の不安障害と分類すべきかどうかを疑問に思っています。これらの障害の神経回路に関するさらなる研究は、この問題の解明に役立つはずです。
全般性不安障害(GAD)に関する最近の研究では、病気に関連した代償性変化の可能性を含め、不安/恐怖回路内の機能的接続における潜在的に重要な変化が浮き彫りになっています。 Amit Etkinらは、健康な対照者の皮質(dIPFCおよび後頭頂皮質)の実行制御ネットワークと扁桃体の間に接続性がほとんどないことを発見した。対照的に、GAD患者は、扁桃体と実行制御領域の間の結合性が強化されているが、島皮質と帯状皮質が関与する顕著性ネットワークへの結合性は低下している。これらの著者らは、皮質制御ネットワークの関与の強化は、感情出力の制御を維持するのに役立つ代償的適応である一方、顕著性システムへの接続性の変化により、自律神経系出力(心拍数、発汗など)の調節に問題が生じる可能性があると推測しています。 。
感情障害: 原発性大うつ病
うつ病障害は、不安障害と同様、さまざまな病気のグループです。私たちはこのことを第 2 章で強調しましたが、脳のネットワークが大うつ病にどのように関与しているかを考えるときに、もう一度強調します。以下の議論で私たちが焦点を当てるのは、他の精神疾患がないにもかかわらず存在する、または明らかに他の疾患の発症に先立って存在する「原発性」非精神性気分障害であり、高度に家族性の型の大うつ病に特に注意が払われています。双極性障害 (I 型) に関連して発生するうつ病についても検討しますが、うつ病と躁状態の両方で精神病症状が現れる傾向を考えると、これは別の形態の病気である可能性があると考えられます。いわゆる「軽度の」うつ病を含むすべてのうつ病をひとまとめにすると、脳のネットワークに関して混乱を招くデータが得られる可能性があるため、これらの考慮事項は重要であると考えられます。
Wayne Drevets、Helen Mayberg らによる研究は、大うつ病性障害 (MDD) における認知、感情、動機付けシステムの関与と、機能的接続の変化が症状と回復にどのように寄与するかを説明する上で重要な役割を果たしています。これらの研究は、大うつ病における機能不全の原因となる可能性のある感情システムの回路を定義するのにも役立ちました。 Drevetsらは、高度に家族性のMDDおよび双極性障害を有するよく特徴付けられた被験者を用いて、眼窩前頭皮質、腹外側前頭前野を含む回路における血流と代謝の増加を発見した。
————————図 5-2
自己言及/記憶/未来
デフォルトモードネットワーク背内側 PFC システム
内側 TL システム AmPFC および PCC ハブ
感情ネットワーク扁桃体
Bed N Stria Terminalis 感情/バイアス
報酬制度
ヌック・アカンベンス、中性尾状腹側淡蒼球
VTA/SN
モチベーション/喜び
内側前頭前野ネットワーク VmPFC、中眼窩 Ctx、吻側島、sgACC
↑
視床内側背筋
統合
応答システム 視床下部 HPA 軸
中水道周囲灰色脳幹核
覚醒/ストレス反応
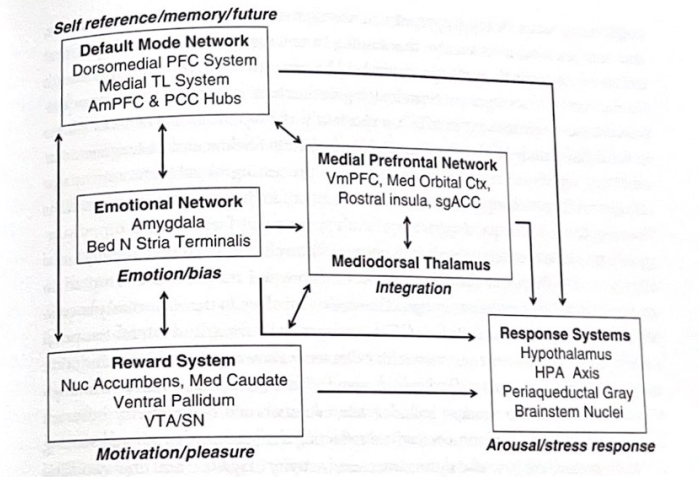
図 5-2 ディプレッション回路。この図は、原発性大うつ病に関与すると考えられる主要な領域とつながりを示しています。この回路は、この複雑な一連の障害における感情、認知、動機付けシステムの役割を浮き彫りにしています。 (Wayne Drevets と Joel Price によって提供された説明を基にしています。) 解剖学的構造の位置については、付録を参照してください。略語: TL (側頭葉)、PCC (後帯状皮質)、sgACC (前帯状皮質下)。
————————
(vIPFC)、および膝下および前帯状皮質 (sgACC および pgACC)。重要な回路には、扁桃体、腹側線条体、内側視床も含まれます。他の研究では、PFC、sgACC、海馬、扁桃体の異常の重要性が強調されています。 MDD に関連する皮質の変化は、より高次の感情体験や認知機能障害に寄与すると考えられていますが、皮質下の変化は感情処理、感情記憶、記憶想起、モチベーション、特に目標を達成するためのモチベーションの変化に寄与すると考えられています。指向のタスク。図 5-2 は、大うつ病に関与する神経回路の概要を示しています。
画像研究では、sgACC と扁桃体の活動の増加が活動性うつ病の症状の重症度に相関することから、sgACC が重要なノードとして浮上しています。対照的に、vIPFC および外側眼窩皮質の変化はうつ病の重症度と負の相関があります。これらの部位間およびこのネットワーク内の相互作用は症状の発生に重要であると考えられており、内側 PFC の活動の低下により扁桃体の脱抑制が生じ、MDD では内分泌系、自律神経系、報酬系、注意系が変化します。興味深いことに、うつ病の治療法が異なれば、コアネットワークに異なる方法で影響を与えるようであり、これはおそらく治療法の基本的なメカニズムを反映していると考えられます。 sgACCを標的とした抗うつ薬と脳深部刺激は、sgACCと扁桃体の過剰活動を減少させるようですが、精神療法的介入(対人関係療法や認知行動療法)はvIPFC、眼窩皮質、前島皮質の活動を増加させるようです。これらの研究から明らかとなったもう 1 つの発見は、pgACC の活性の増加は治療に対する反応の改善を予測する可能性があるということですが、その理由はまだ明らかではありません。画像研究では、MDD では海馬の変化が顕著であることもわかっています。重要なのは、海馬がデフォルトモードのICNの重要な役割を担っており、Yvette Shelineらによる最近の研究で、大うつ病患者のデフォルトモード処理に大きな変化が見られることが実証されたということである。これらの変化には、海馬だけでなく、海馬に栄養を与える扁桃体や海馬傍皮質の活動の増加も含まれます。これらの後者の研究では、デフォルトネットワークは、うつ病の人がネガティブな内容のタスクに従事しようとしたときに、刺激誘発性の活動が強化され、下方制御されないことを示しました。異常な処理に関与する領域には、腹内側 PFC、ACC、外側頭頂葉皮質、および外側側頭葉皮質が含まれていました。これらの研究は、視覚要求の高いタスクに注意を集中させるよう求められたうつ病患者の脳機能と接続性の変化を示す他の研究と一致しています。これらの変化には、前頭葉皮質、頭頂葉皮質、および視覚皮質間の機能的接続の変化が含まれており、おそらく注意/再配向 ICNS の変化を反映しています。うつ病の被験者は、課題遂行に必要な注意要求によって調節されるsgACCおよび内側眼窩皮質の活動の増加も示した。 19件の研究を対象とした最近のメタ分析では、治療反応の予測因子として吻側ACC活性の増加の重要性が強調されており、この領域の活性が低いことが、デフォルトネットワークを非活性化してタスク活性化ネットワークをオンラインにすることの困難に寄与している可能性があると推測されている。
大うつ病の機能回路に関与する多くの領域も構造変化を示します。これについては次の章で詳しく説明します。現時点では、海馬、扁桃体、sgACC、PFC の他の領域、および大脳皮質の側面にも構造変化を示す証拠があることに留意することが重要です。これらの変化がどのように起こるかは明らかではありませんが、うつ病患者の中には病気の初期に変化を示し、おそらく病気の発症よりも前に変化を示す人もいれば、病気の期間と相関すると思われる変化を示す人もいます。構造変化は、さまざまな地域の特定の細胞集団の変化にも関連付けられています。例えば、sgACC および扁桃体ではグリア細胞の損失、特に扁桃体の希突起膠細胞の損失が見られます。希突起膠細胞は、あるニューロンから別のニューロンに情報を伝える軸索のミエリン絶縁体を形成するのに役立ちます。また、脳の領域をつなぐ白質路の形成にも役立ちます。 sgACCでは、Dost Öngürらは、MDD患者ではグリア細胞が20%以上減少したが、双極性障害患者では約40%減少したと報告した。他の研究では、皮質の他の領域におけるグリア細胞および神経細胞の喪失が示されています。特に双極性障害では、海馬などの一部の領域の介在ニューロンにも変化が見られます。 Francine Benesらは、双極性障害患者のCA2領域の非錐体ニューロンが約40%減少し、CA3領域の細胞数が減少する傾向があることを報告した。心理社会的ストレスはシナプスの発達と維持に影響を及ぼし、ニューロンの樹状突起の縮小を引き起こすことが知られています。樹状突起は次の場所にあります。
どのニューロンが通常他のニューロンから入力を受け取るか。これらの変化は細胞の喪失と共謀して、重度の気分障害を患う被験者で観察される局所的な脳容積の減少を引き起こす可能性があります。特定の領域での変更は重要であり、それらの領域での処理欠陥の原因となる可能性がありますが、特定の領域での変更が単独で発生するわけではなく、ネットワーク全体のパフォーマンスに影響を与えることを考慮することが重要です。精神科の症状や障害の中心にあるのはネットワーク機能不全です。
最後に、感情処理は、他の多くの脳計算と同様に、半球の側性化を示すことを考慮することも重要です。これは、前頭葉に影響を与える脳卒中を患っている人に最もはっきりと観察されます。ここで、左前頭部の損傷は通常、抑うつ気分と関連していますが、右前頭部の脳卒中は、一部の被験者では気分の変化がない、または躁状態のような症状と関連しています。この観察は、覚醒と生存を監視する自律神経系の一部である交感神経系からの情報は右半球で優先的に処理されるのに対し、弛緩と親近感を扱う副交感神経系からの入力は右半球で処理されるという事実を反映している可能性があります。左半球。前島皮質および関連する吻側 ACC も、自律神経処理および気分状態に関連する主観的感情の生成に重要な役割を果たしています。左側の脳卒中の後、右半球が過剰に活動する可能性があり、これがネガティブな気分状態を引き起こす可能性があります。
ここで概説した研究は、大うつ病性障害を感情、認知、または動機の主要な欠陥として分類するという点で複雑であることを示しています。感情的な ICNS がこれらの障害の中心であるように見えますが、顕著な認知、注意、および動機付けの欠陥も伴います。うつ病に関連する認知の変化は、障害を引き起こす上で特に重要である可能性があります。 Luke Clarkらは、前述のPFC皮質下回路内の機能不全に起因するうつ病患者の実行機能、記憶、感情処理、フィードバック感度の問題を強調した。したがって、大うつ病性障害は、心の 3 つの側面すべてを統合するために必要なシステムの障害を表している可能性があります。大きな疑問は、複数の脳ネットワークにわたる接続と機能の変化がどのように起こるかに関するものである。答えは不完全ですが、高度に接続され、高度に可塑性のある神経ハブが重要な役割を果たす可能性があるようです。たとえば、Shelineらは最近、感情、認知制御、およびデフォルトモードのニューラルネットワークが、うつ病患者の両側背側内側PFCの同じ領域への安静状態の接続性の増加を示していることを観察した。彼らは、背側内側 PFC のこの領域を「背側結合」と呼んでいます。この背側結合は、病気の際にネットワークを結び付ける役割を果たしているようで、背側結合と 3 つのネットワークの接続の程度は疾患の重症度と相関します。他の最近の研究では、重要な前脳領域と中脳領域を結び付ける外側手綱核などの領域が、脳に関連する多くの行動の調節に重要な役割を果たしていることが示唆されています。したがって、複数のノードがうつ病の根底にある回路を統合することに関与している可能性があり、これらのノードは、標的を絞った治療介入の重要な領域となる可能性がある。
動機付け障害: 薬物乱用
すべてではないにしても、ほぼすべての依存性薬物が同じ神経系を攻撃するため、薬物乱用障害の神経回路と細胞機構については、他のどのグループの精神疾患よりも多くのことが知られている可能性があります。これらの疾患に関与する主要なネットワークは、げっ歯類とヒトで同様に組織されています。この類似性により、詳細に特徴付けられた動物モデルの開発や、細胞、シナプス、およびシステムレベルでの変化の詳細を解明するための高度な生理学的および分子ツールの使用が可能になりました。エリック・ネスラー、スティーブ・ハイマン、ジョージ・クーブ、ロブ・マレンカ、ピーター・カリバス、ノラ・ヴォルコウ、リード・モンタギューなどを含む多くの科学者がこの研究で重要な役割を果たし、その結果は考え方を理解するためのモデルを提供しました。他の精神疾患の生物学について。
我々が指摘したように、依存性薬物は、中脳、腹側被蓋野(VTA)、中脳辺縁系および中皮質ドーパミン系の起源を含むネットワークにおける神経伝達物質ドーパミンの作用を調節する(図5-3)。 VTA のニューロンは、側坐核、海馬、PFC、その他の領域に投影されます。 VTA-側坐筋-PFCネットワークは、乱用薬物の急性効果に関与する主要な回路であり、私たちの根底にあると考えられている回路でもあります。
————————図 5-3
モチベーション・やりがい
感情/感情
認知/記憶制御
側坐核腹部淡蒼球 VTA/SN
扁桃体床と終端線条 視床下部 脳幹核
↓
海馬島 前頭前皮質
酩酊 暴飲暴食
気分の落ち込み/不安の離脱
こだわり 認知欠陥 依存症
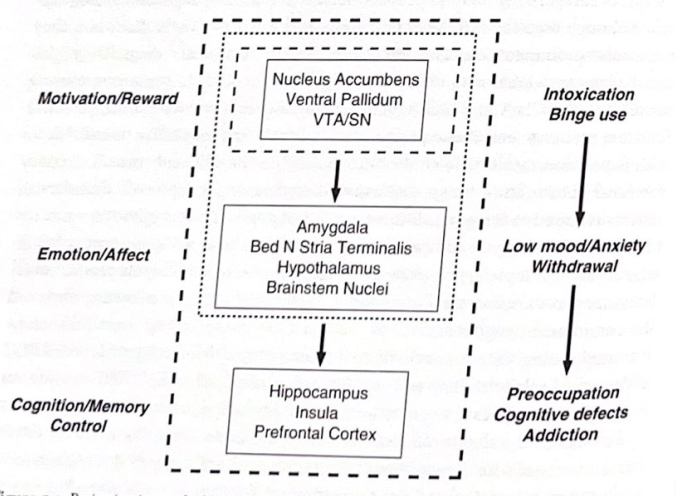
図 5-3 脳の回路と依存症。この図は、依存性薬物の影響に関与する神経回路と、依存症が進行するにつれて回路がどのように複雑になるかを示しています。 (George Koob と Nora Volkow によって提供された説明を基にしています。) 解剖学的構造の位置については、付録を参照してください。
————————
モチベーションと報酬システム。このネットワークでは、ドーパミン ニューロンの発火が、経験が期待よりも良いか悪いかを示す報酬予測信号として機能すると考えられます。期待を超える経験はこれらのニューロンの発火を促進しますが、期待を下回る経験は発火を低下させます。ドーパミン ニューロンは継続的に側坐核と PFC に情報を送信し、その発火率の変化によりシナプス後構造のドーパミン レベルが増加または減少し、リード モンタギューの言葉を借りれば、ドーパミンが「評価」シグナルとして機能するようになります。
ドーパミンは古典的な低速伝達物質であり (第 3 章を参照)、脳領域内および脳領域全体の活動を組織化し、調整するのに役立ちます。また、これらの領域の入力と出力の調整にも関与します。側坐核では、ドーパミン信号が価値判断に影響を与え、何かに近づくべきか避けるべきかの決定に役立ちます。側坐核内のさまざまな回路が、接近と回避の決定に役割を果たしている可能性があります。腹側淡蒼球および背側線条体-淡蒼球系を介した側坐核の出力は、運動反応の調整に役立ちます。 PFC では、ドーパミンはニューロンがどの信号に注意を払うかを決定するのに役立ち、事実上、入力の「顕著性」を決定するのに役立つ一種のゲート機能を実行します。
VTA-側坐核-PFC ループは、薬物乱用に関与する最も単純化された回路を表します。扁桃体 (およびその接続部) や海馬 (およびそれに関連する ICN) など、他の領域も寄与しています。これらの関係と、これらの領域およびネットワークに対するドーパミンの影響は、薬物乱用障害における学習および記憶だけでなく感情システムの関与への経路を提供します。乱用薬物の作用に関与する回路の研究により、扁桃体本体、側坐核の殻(外側)領域、およびBNSTを含む扁桃体の分布した解剖学的構造と接続性が明らかになりました。乱用された薬物のほとんどは、これらの領域の 1 つまたは複数に直接影響を与えます。 PFC 内のいくつかの地域も、この拡張ネットワークに関与しています。皮質領域には、目標に向けた意思決定に関与し、おそらく価値信号の解釈に関与する腹内側 PFC と、作業記憶、感情制御、および認知制御に重要な dIPFC が含まれます。 ACC も関与しており、薬物乱用障害を持つ人ではこの領域の活性が低下しており、パフォーマンスの監視や認知制御に問題が生じています。物質乱用障害における高次脳中枢の関与により、ノラ・ボルコウらはこれらの障害を単なる衝動制御や動機付けの欠陥ではなく「神経認知」の問題と呼ぶようになった。これは深遠な声明であり、私たちが強調したいことは精神医学全体に貫かれています。本格的な主要な精神疾患はすべて「神経認知」障害です。
一般的な用語で言えば、ほとんどの中毒性のある薬物は脳のモチベーション回路を「ハイジャック」し、事実上、自然な報酬を置き換えます。これらの薬剤は、いくつかのメカニズムを使用して VTA 末端からのドーパミンの放出を促進することによってこの偉業を達成します。コカインとアンフェタミンは、ドーパミンの取り込みをブロックする(コカイン)か、ヘテロ交換を介してドーパミンの放出を促進する(アンフェタミン)ことにより、側坐核やその他の領域のドーパミン輸送体の機能を変化させます。
後者の点は、ドーパミンが放出されるのと引き換えに、アンフェタミンがドーパミン輸送体によって取り込まれることを意味します。他の薬物(アヘン剤など)は、VTA のドーパミン ニューロンの発火を変化させることによってドーパミン レベルを上昇させます。
ドーパミンに対する依存性薬物の重大な影響のため、他の「依存性」行動がドーパミンシステムに影響を与えるかどうかを決定することに関心が集まっています。たとえば、ドーパミン系の変化は、病的なギャンブル、強迫的な買い物、おそらくは強迫的なセックスや過食など、特定の人間の「衝動制御障害」で発生する可能性があります。同様に、神経画像研究では、特定のビデオ ゲームをプレイするとドーパミンの放出が促進されることが示唆されており、これらのゲームが「中毒性」があるのではないかという考えに寄与しています。黒質におけるドーパミンニューロンの喪失に起因する障害であるパーキンソン病の治療に使用されるものを含む、ドーパミン系に影響を与える治療薬も、上記の衝動制御障害の一部を含む依存性行動を引き起こす可能性があります。これらの発見は、パーキンソン病の複雑さと、多数の動機付けられた行動の調節におけるドーパミンの役割についての理解の進展に貢献しました。重要なのは、乱用薬物はドーパミンレベルを上昇させるのに特に優れているため、他の行動よりもはるかに効果的であり、依存性がより高いということです。
ドーパミン放出に対する依存性薬物の影響は、薬物が脳内の標的に到達するとすぐに現れます。しかし、薬物乱用障害は慢性的な問題であり、最も重篤な状態になるには薬物への繰り返しの曝露が必要です。このため、セカンドメッセンジャー、遺伝子発現、タンパク質合成への影響など、これらの薬物によって誘発されるVTA-側坐筋系の長期適応を調べる研究が促進されています。側坐核では、ドーパミンが D1 型受容体に結合し、GTP 結合タンパク質 (G タンパク質) を介してアデニル酸シクラーゼを活性化します。これは、セカンドメッセンジャーであるサイクリックAMPの合成につながります。次に、サイクリック AMP は、CREB (サイクリック AMP 応答エレメント結合タンパク質) を含むさまざまなタンパク質をリン酸化する酵素であるプロテイン キナーゼ A を活性化します。 CREB は、ドーパミンによる遺伝子発現の変化を可能にする転写因子です。 CREB は、他の遺伝子の発現を駆動する転写因子でもある前初期遺伝子 (例: c-fos、fos-B) の活性化を促進します。この一連の出来事は、ニューロンの機能と構造に長期的な影響をもたらし、これらの変化は、個々のニューロンとネットワークに対する乱用薬物の長期的な影響を促進するために重要である可能性があります。依存性薬物は、第 6 章で説明するシナプス可塑性メカニズムを利用して、VTA とその接続領域のシナプスの機能に大きな影響を与えます。VTA のドーパミン ニューロンは、PFC、視床下部、および被蓋核から興奮性グルタミン酸入力を受け取ります。説得力のある証拠は、これらの興奮性シナプスが乱用薬物への曝露後に長期増強(LTP)を示すことを示しています。 VTA LTP は、薬物に対する初期の行動反応にとって重要であり、ドーパミン ニューロンの出力増加につながり、コカイン、アンフェタミン、ニコチン、エタノールなどのさまざまな薬物を投与した後の生きた動物で観察されます。この持続的なシナプス増強には、海馬における LTP (第 6 章) と同様、N-メチル-D-アスパラギン酸 (NMDA) 型グルタミン酸受容体の活性化が関与します。興味深いことに、一部の薬物 (コカインやエタノールなど) は、生体内で 1 回曝露されただけでげっ歯類の VTA で LTP を生成します。これは、長期にわたるプロセスを開始するには 1 回の薬物曝露だけで十分であることを示しています。他の薬(抗うつ薬など)にはこのような効果はありません。 VTA の初期の LTP は、薬物体験に動機付けの重要性を付加する役割を果たし、このシステム内の機能の長期的な変化をさらに促進するのに役立つ学習形式をもたらすと考えられています。
シナプスの変化は、VTA ニューロンの主な標的である側坐核でも起こります。側坐核には、背側線条体 (尾状被殻) の延長部であるドーパミンである「コア」領域と、延長された扁桃体の一部である「シェル」領域の 2 つの主要な部分があります。 LTP に加えて、長期シナプス可塑性の別の形態である長期シナプス抑制 (LTD) があり、これはシナプス伝達の持続的な減少をもたらします。重要なことに、薬物曝露は側坐核のシェル領域にグルタミン酸シナプスのLTDを生成しますが、コア領域には生成しません。この LTD は、乱用薬物に対する行動的感作と相関しているようです。依存症ではない動物は時間の経過とともに側坐核の急性LTDから回復するが、持続的な「依存症様」行動(薬物を自己投与する傾向を意味する)を示す動物は永続的なシナプス変化を示す。乱用薬物は側坐核の細胞外グルタミン酸レベルの長期変化にも関連しており、これらの変化はグリア細胞内のグルタミン酸トランスポーターの一種(シスチングルタミン酸トランスポーターと呼ばれる)に対する影響によって媒介されるようです。
げっ歯類における高度な細胞および分子研究と人間における機能画像研究を組み合わせた結果、依存性障害を「神経認知」の観点からどのように概念化できるかについての洞察が得られました。 George Koob と Nora Volkow が記述したモデルでは、薬物中毒は 3 つの段階を経て進行すると見られています (表 5-4)。最初は、薬物の過剰摂取と中毒が繰り返される段階があり、すでに説明した VTA 側坐筋回路が強く関与します。薬物使用を続けると、薬物中毒と薬物離脱が繰り返され、それに伴う否定的な気分や不安が生じる段階に入ります。この段階では、VTA側坐骨の外側の回路が拡張され、拡張された扁桃体とその感情調節における役割が関与します。
薬物使用がさらに長期化すると、個人は薬物に夢中になり、薬物使用に対する期待が高まる段階に入り、認知とより高いレベルの制御の基礎となる回路を含む、さらに分散されたニューラルネットワークが関与します。この第 3 段階の主要な構造には、眼窩前頭皮質、背側線条体、PFC、海馬、および島皮が含まれます。これらは、薬物渇望や薬物乱用の文脈的側面、さらには記憶機能障害に関与していると考えられる拡張されたネットワークです。薬物使用に
おいては状況が重要な役割を果たしており、効果的な治療法には依存症者の生活の再構築が必要となる理由の 1 つとなっています。薬物乱用の第 3 段階に関与する他の領域には、ACC、下前頭皮質、抑制制御、注意、作業記憶に重要な回路である dIPFC などがあります。
————————
表 5-4 依存症の 3 つの段階
暴飲暴食と酩酊
VTA-側坐核「報酬」回路が関与
度重なる離脱と悪影響
拡張した扁桃体(終端線の床核を含む)も関与します。
こだわり、習慣、実行機能障害
海馬、眼窩前頭皮質、PFC、前島、背側線条体も関与します。
————————
重要なのは、薬物の使用と離脱の繰り返しサイクルに関連するシナプス可塑性が、この回路の時間の経過とともに拡大する鍵であり、相互接続されたネットワークの機能不全を引き起こし、心のあらゆる側面に悪影響を与えることです。時間の経過と病気の繰り返しによる機能不全の回路の同様の拡大が、他の精神疾患の病因と複雑さに寄与する可能性があると考えるのは、あまり大きな推定ではありません。薬物乱用の回路を研究する利点は、障害が特定の回路から始まり、詳細に定義して研究できることです。病気が進行すると、神経ネットワークがさらに拡大し、より複雑になります。図 5-3 は、薬物中毒の段階に関与すると考えられる神経回路の概要を示しています。
この話の多くの側面は不確かなままですが、重要な点は、動機付けシステムがどのように機能するか、そして乱用薬物がどのようにシステムを攻撃して急性および長期的な影響を生み出すかについて、現在では多くのことがわかっているということです。この情報は、薬物乱用症候群の新しい治療戦略を開発するためのさまざまなアイデアを刺激しました。例えば、上記のシスチン-グルタミン酸トランスポーターが細胞外グルタミン酸レベルを変化させることにより治療薬理学の標的として機能するかどうか、またトランスポーターを介した細胞外グルタミン酸レベルの変化がグルタミン酸の特定のサブタイプに対するグルタミン酸の作用に影響を与えるかどうかを決定することに関心が寄せられている。受容体(特に代謝指向性受容体)。これらの受容体は、治療介入の標的となる可能性があります。
中毒性薬物のモチベーションシステムに対する影響は、細胞レベルおよびシナプスレベルで長期間持続します。したがって、物質乱用症候群が慢性的かつ再発性の機能障害を伴うことが多いのは驚くべきことではありません。扁桃体と海馬の関与により、これらの症候群に感情的および学習的要素が加わり、薬物の慢性的な影響がさらに悪化します。依存症の分野での研究は、動機、感情、認知システムがどのように絡み合っているのか、そして病気の進行に伴ってこれらの神経システムがどのように複雑な回路に関与するのかを理解することにも影響を及ぼします。
その他の乱用薬物: 幻覚剤
上で概説した研究には、主にドーパミン系を攻撃する依存性薬物が含まれています。別のグループの乱用薬物は異なる作用機序を持ち、知覚と認知を変える能力を反映して「幻覚剤」と呼ばれています。これらの薬物は、その主な効果が感覚処理に影響を与えると思われるため興味深いものであり、おそらく高次の知覚処理に関与するメカニズムについての洞察を提供します。ここでは、幻覚剤の 2 つのクラスについて簡単に説明します。LSD (リセルグ酸ジエチルアミド) とシロシビン (「キノコ」) を含むグループと、MDMA (3,4-メチレンジオキシメタンフェタミン、別名「エクスタシー」) に代表されるクラスです。これらの薬物は乱用されていますが、伝統的な意味での中毒性はありません。これらの幻覚剤に対する身体の耐性は非常に急速に発達するため、幻覚剤は時折使用される傾向があり、依存性薬物に見られるような渇望、強迫的な薬物探索、痛みを伴う禁断症状のサイクルには通常は至りません。興味深いことに、幻覚剤は精神機能に悪影響を及ぼしますが、主に中枢の動機付けシステムに影響を与えるわけではありません。
LSD とシロシビンは、時間の歪み、視覚的感覚イメージ (錯覚を含む)、および場合によっては共感覚を引き起こす可能性があります。共感覚とは、感覚が絡み合う現象を指します(たとえば、色が聞こえたり、音が見えたりする場合があります)。使用者らは、これらの経験の一部が深い自己超越、神秘的な洞察、知識につながると指摘していますが、これらの薬物がもたらす可能性のある深刻な副作用を考慮すると、これは事実というよりも逸話にすぎません。 David Nicholsらは、LSD型幻覚剤が、Gタンパク質の特定のサブタイプを介して細胞内シグナル伝達経路に結合しているセロトニンSHT2A受容体に対するアゴニストとして作用することを示した。 Javier González-Maesoらは、非幻覚性SHT2Aアゴニストではなく幻覚剤が、独特のシグナル伝達特性を持つ5HT2A受容体サブタイプを介して特定の皮質ニューロンに直接影響を与えることによって作用する可能性があることを実証することで、これらの発見を拡張した。これらの発見は、薬物がセロトニンなどの分散伝達系に影響を与える場合でも、受容体の分子サブタイプやシグナル伝達カスケードを介して高度に特異的な効果が達成される可能性があることを示唆している。これにより、特定の皮質ニューロンおよび領域の操作がどのようにネットワーク特性に影響を与え、感覚体験の変化をもたらすかを理解するための扉が開かれます。
MDMA (エクスタシー) は、攻撃性を低下させるとともに、警戒心と共感、思いやり、親密さの感情を高めることが報告されています。この薬剤は、ホルモン/伝達物質であるオキシトシンだけでなく、セロトニン作動性およびアドレナリン作動性システムにも影響を与えます。オキシトシン系は交尾および社会的結合行動に役割を果たしているようであり、この薬物の研究により社会的相互作用に関与する脳システムについての洞察が得られる可能性がある。サイロシビンとMDMAは、末期がん患者の不安に対する治療法の可能性として研究されています。このような臨床使用は、MDMA への慢性的な曝露がセロトニン系に有毒な影響を与える可能性があるため、複雑になります。
要約: 繰り返すテーマ
機能的神経画像研究は、さまざまな精神疾患に関与する脳領域間の重複を説得力を持って実証しました。より明らかになりつつあるのは、特定の脳領域が特定の処理タスクにどのように寄与しているのか、そして特定の領域の機能不全がICNまたは他のICNSの残りのノードの機能にどのように重大な影響を与える可能性があるのかということです。ネットワークの動作、つまり入力をどのように処理するか、ローカルで情報を処理する方法、およびその情報を使用して他の地域と通信する方法を理解することは、精神科の症状や障害を理解するために重要です。一部の脳領域は、その高い接続性により、複数の回路に関与しています。したがって、複数の病気にわたってこれらの領域の機能変化が見つかることは予期せぬことではありません。海馬、扁桃体、側坐核、PFC などがそのような領域の明確な例です。認知、感情、動機の基礎となるこれらの共有回路内で疾患の特異性がどのようにして生じるのかは必ずしも明らかではありませんが、特定の疾患の初期に関与する主要なノードまたはネットワークが、その臨床疾患の性質を定義する上で大きな役割を果たしているのではないかと私たちは推測しています。たとえば、依存性薬物は明らかに早期に動機/報酬システムを攻撃するため、この種の処理の欠陥がこれらの障害の主な問題になります。また、これらの回路とさまざまな病気の間には非常に多くの重複があるため、特に病気が持続し、ネットワーク全体に病気による可塑性が生じる可能性を考慮すると、病気の実質的な「併存症」が存在することは驚くべきことではない、とも主張したいと思います。進捗。繰り返しになりますが、心の 3 つの側面すべてがすべての障害に関与しています。この事実は併存疾患を保証します。これを整理する鍵は、特定の病気でどの処理欠陥が最も顕著であるのか、そして最終的には何がそれらのICNをシナプス、細胞、分子レベルで脆弱にするのかを理解することにあるようです。また、病気の経過の中でどの脳の変化が最も早く起こり、どの変化が二次的、三次的、または代償的であるかを知ることも重要です。しかし、代償性変化は必ずしも有益ではなく、最終的には病気の表現型に寄与する可能性があります。以前に述べたように、細胞およびシナプスの変化は薬理学的介入の潜在的な標的である一方、ICN/システムレベルでの機能の変化は精神療法およびリハビリテーション戦略の潜在的な標的です。ほとんどの患者で最適な臨床結果を得るには、両方のタイプの介入が必要になる可能性があります。
過度に単純化する危険を承知で、この章で概説したさまざまな脳領域が情報処理と精神機能障害に寄与する可能性があることを説明して、この議論を終了します。この議論は、トーマス・ミークスとディリップ・ジェステによる「知恵の神経生物学」に関する最近の論文に基づいています。新皮質、特に PFC は、脳内で最高レベルの処理と最高レベルの抽象化を提供します。側方およびdIPFCは、作業記憶と、感情や衝動に対する意識的なトップダウン制御の認知的側面にとって重要です。これらの領域は、私たちの脳が直面する繰り返しの課題の 1 つである曖昧さへの対処においても重要な役割を果たします。内側および腹内側の PFC は、自己反省と意思決定の感情的な要素に重要ですが、眼窩前頭皮質は、反応抑制を介して衝動性を制御し、刺激と報酬に感情的な価値を付加する役割を果たします。頭頂皮質の一部、特に頭頂前頭結合は、注意を集中させたり、反応の方向を変えたり、知性の基礎となる多くの高次の認知プロセスに重要であるのに対し、上側頭溝の領域は自分と他人、そして他人とを区別するために重要であると考えられています。社会的関連性を判断するため。皮質下の構造は感情と動機に関与しています。扁桃体は、ほぼすべての感情の処理と、物事に対する私たちの反応の決定において重要な役割を果たします。皮質および皮質下への幅広い接続性が、複数の機能の調節、曖昧さの処理、顕著性と重要性の決定における重要な役割の基礎となっている可能性があります。側坐核/線条体系は目標の選択と動機付けに重要であり、報酬の価数と強化を決定するのに役立ちます。 PFCおよび頭頂皮質の皮質下と高中枢の間のどこか、前帯状皮質などの領域は矛盾を検出し、矛盾に対処する回路を活性化しているようです。後帯状皮質は、自己関連(内受容)刺激を決定するのに役立ち、島皮質とともに、おそらく嫌悪感を介して道徳的判断の感覚を提供するのに重要である可能性があります。重要なのは、これらの領域はいずれも単独で機能するわけではなく、各領域によって提供される計算は、脳の健全な機能に不可欠なネットワーク内およびネットワーク間の処理に寄与するということです。病気は、特定の主要な領域、特に領域の組み合わせの欠陥によって、あるいはこれらのネットワーク内の接続の変化によって発生する可能性があります。最終的に、多くの脳機能の目標は、原始中枢 (扁桃体、VTA 側坐骨、視床下部/脳幹) を秘密に保つことであると考えられます。
————————
留意事項
精神障害は、脳ネットワークの構造と機能が関与する複雑な問題です。伝達システムの変化が障害の一因となっている一方で、病気は単に「化学的不均衡」の結果ではありません。
特定の ICN の役割に関する作業はまだ初期段階にあります。それにもかかわらず、このレベルの分析は精神障害の症状と表現型を定義する上で多大な可能性を秘めており、より有効な診断アプローチにつながる可能性があります。 ICN レベルの分析は、より優れた心理療法およびリハビリテーション戦略の開発に貢献する可能性もあります。
恐怖と薬物乱用の十分に特徴付けられた動物モデルが存在するため、これらの状況において細胞とシナプスの変化がどのように共謀してネットワーク機能の変化を引き起こし、ひいてはネットワーク機能の変化がどのように行動の変化をもたらすかについての理解がさらに進んでいます。 。動物実験での発見は、不安症や薬物乱用障害を薬理学的および心理療法レベルで治療する方法についての新しいアイデアにつながっています。
————————
推奨される読み物
Bassett, D.S.、Bullmore, E.、Verchinski, B.A.、Mattay, V.S.、Weinberger, D.R.、および Meyer-Lindenberg, A. (2008)。健康と統合失調症における人間の皮質ネットワークの階層的組織。神経科学ジャーナル、28、9239-9248。
Davis, M.、Walker, D.L.、Miles, L.、Grillon, C. (2010)。ラットとヒトにおける一時的恐怖と持続的恐怖: 恐怖と不安における拡張扁桃体の役割。神経精神薬理学 レビュー、35、105-135。
W.C. ドレベッツ、J.L. プライス、M.L. フューリー (2008)。気分障害における脳の構造的および機能的異常: うつ病の神経回路への影響。脳の構造と機能、 213,93-118。
Koob、GF、Volkow、ND (2010)。依存症の神経回路。神経精神薬理学レビュー、35、217-238。
LeDoux, J. (2002)。シナプス自己: 私たちの脳がどのようにして私たちになるのか。ニューヨーク:バイキング・プレス。ルイス D.A.、スウィート R.A. (2009)。神経回路の観点から見た統合失調症: 合理的な薬物療法への進歩。臨床研究ジャーナル、119、706-716。
D. ナット、L.A. キング、W. ソールズベリー、C. ブレイクモア (2007)。潜在的な誤用の薬物の害を評価するための合理的な尺度の開発。ランセット、369、1047–1053。
その他の参考文献
ベネス、F.M. (2007)。神経回路内の統合失調症および双極性障害の固有のエンドフェノタイプとその分子調節機構の探索。統合失調症速報、33、932-936。
Bentall, R. P.、Rowse, G.、Shryane, N.、Kinderman, P.、Howard, R.、Blackwood, N.、他。 (2009年)。偏執的な妄想の認知的および感情的構造。一般精神医学のアーカイブ、66,236-247。
Bloch, M. H.、Landeros-Weisenberger, A.、Rosario, M. C.、Pittenger, C.、および Leckman, J. F. (2008)。強迫性障害の症状構造のメタ分析。アメリカ精神医学ジャーナル、165、1532-1542。
Buckner, R.L.、Andrews-Hanna, J.R.、Schacter, D.L. (2008)。脳のデフォルト ネットワーク: 解剖学、機能、病気との関連性。ニューヨーク科学アカデミー年報、1124、1-38。
ブルモア、E.、スポンズ、O. (2009)。複雑な脳ネットワーク: 構造および機能システムのグラフ理論分析。 Nature Reviews Neuroscience、10、186-198。
Cavedini, P.、Zorzi, C.、Piccinni, M.、Cavallini, M. C.、および Bellodi, L. (2010)。強迫性患者とその影響を受けていない親族における実行機能障害: 新しい中間表現型の探索。生物学的精神医学、67、1178-1184。
Chamberlain, S.R.、Menzies, L.、Hampshire, A.、Suckling, J.、Fineberg, N.A.、del Campo, N.、他。 (2008年)。強迫性障害患者およびその影響を受けていない親族における眼窩前頭葉機能障害。サイエンス、321、421-422。
Clark, L.、Chamberlain, S.R.、および Sahakian, B.J. (2009)。うつ病における神経認知メカニズム: 治療への影響。神経科学年次レビュー、32、57-74。
Cloninger、C.R. (2004)。良い気分: 幸福の科学。ニューヨーク:オックスフォード大学出版局。
Dagher、A.、Robbins、T.W. (2009)。性格、依存症、ドーパミン: パーキンソン病からの洞察。ニューロン、61、502-510。
Desseilles, M.、Balteau, E.、Sterpeních, V.、Dang-Vu, T. T.、Darsaud, A.、Vandewalle, G.、他。 (2009年)。うつ病における無関係な視覚情報の神経フィルタリングの異常。神経科学ジャーナル、29、1395-1403。
デビンスキー、O. (2009)。妄想上の誤認と重複。右脳障害、左脳妄想。神経学、72、80-87。
アイゼンバーグ、D.P.、バーマン、K.F. (2010)。統合失調症における実行機能、神経回路および遺伝的メカニズム。神経精神薬理学レビュー、35、258-277。 Etkin, A.、Prater, K.E.、Schatzberg, A.F.、Menon, V.、Greicius, M.D. (2009)。全般性不安障害における扁桃体小領域の機能的接続の破壊と代償ネットワークの証拠。一般精神医学アーカイブ、66、1361-1372。
Fineberg, N.A.、Potenza, M.N.、Chamberlain, S.R.、Berlin, H.A.、Menzies, L.、Bechara, A.、他(2010年)。動物モデルからエンドフェノタイプまで、強迫的および衝動的な行動を調査する: 物語的なレビュー。神経精神薬理学、35、591-604。
フレッチャー、P.C.、フリス、C.D. (2009)。知覚は信じることです: 統合失調症の陽性症状を説明するためのベイズ的アプローチ。 Nature Reviews Neuroscience、10、48-58。 Giacobbe, P.、Mayberg, H.S.、および Lozano, A.M. (2009)。脳の恒常性維持機構の不全としての治療抵抗性うつ病: 脳深部刺激の意味。実験神経学、219、44-52。
González-Maeso, J.、Weisstaub, N.V.、Zhou, M.、Chan, P.、Ivic, L.、Ang, R.、他(2007)。幻覚剤は、特定の皮質 5-HT 受容体媒介シグナル伝達経路を動員して、 行動に影響を与える。ニューロン、53、439-452。
Hare, T.A.、Camerer, C.F.、Rangel, A. (2009)。意思決定における自己制御には、vmPFC 評価システムの調整が含まれます。サイエンス、324、646-648。
Harrison, B. J.、Soriano-Mas, C.、Pujol, J.、Ortiz, H.、Lopez, M.、Hernandez-Ribas, R.、他(2009年)。強迫性障害における皮質線条体の機能的接続の変化。一般精神医学アーカイブ、66、1189-1200。
Heller, A.S.、Johnstone, T.、Schackman, A.J.、Light, S.N.、Peterson, M.J.、Kolden, G.G. 他(2010年)。大うつ病におけるポジティブな感情を維持する能力の低下は、前頭線条体脳の活性化維持の低下を反映している。米国科学アカデミー紀要 (米国)、106、22445-22350。
彦坂千里 (2010)手綱核:ストレス回避から価値観に基づく意思決定まで。 Nature Reviews Neuroscience、11、503-513。
ハイマン、S.E.、マレンカ、R.C. (2001)。依存症と脳:強迫とその持続に関する神経生物学。 Nature Reviews Neuroscience、2、695-703。
カリバス、PW (2009)。依存症のグルタミン酸ホメオスタシス仮説。 Nature Reviews Neuroscience、10、561-572。
カプール、S. (2003)。異常な顕著性の状態としての精神病: 統合失調症における生物学、現象学、薬理学を結びつける枠組み。アメリカ精神医学ジャーナル、160、13-23。 Kasanetz, F.、Deroche-Gamonet, V.、Berson, N.、Balado, E.、Lafourcade, M.、Manzoni, O.、他。 (2010年)。依存症への移行は、シナプス可塑性の持続的な障害と関連しています。科学、328、1709-1712。
カウアー、J.A.、マレンカ、R.C. (2007)。シナプス可塑性と依存症。 Nature Reviews Neuroscience、8、844-858。
W. H. ケイ、J. L. ファッジ、M. パウルス (2009)。神経性食欲不振症の症状と神経回路機能に関する新たな洞察。 Nature Reviews Neuroscience、10、573-584。
Lahti, A.C.、Weiler, M.A.、Holcomb, H.H.、Tamminga, C.A.、および Cropsey, K.L. (2009)。大脳辺縁系回路の調節は抗精神病薬に対する治療反応を予測する:統合失調症における機能画像研究。神経精神薬理学、34、2675-2690。 Lesh, T.A.、Niendam, T.A.、M.J. ミンゼンバーグ、C.S. カーター (2011)。統合失調症における認知制御障害:メカニズムと意味。神経精神薬理学レビュー、36、316-338。
Lisman、J.E.、Coyle、J.T.、Green、R.W.、Javitt、D.C.、Benes、F.M.、Heckers、S.、他。 (2008年)。神経伝達物質とリスク遺伝子の相互作用を理解するための回路ベースのフレームワーク 統合失調症。神経科学の動向、31、234-242。
Mansouri, F.A.、Tanaka, K.、Buckley, M.J. (2009)。葛藤によって引き起こされる行動の調整: 前頭前野の実行機能への手がかり。 Nature Reviews Neuroscience、10、141-152。
Mataix-Cols, D.、Wooderson, S.、Lawrence, N.、Brammer, M. J.、Speckens, A.、および Phillips, M. L. (2004)。強迫性障害における、洗濯、確認、溜め込みの症状次元の独特の神経相関。一般精神医学アーカイブ、61、564-576。ミークス、T.W.、ジェステ、D.V. (2009)。知恵の神経生物学: 文献の概要。一般精神医学アーカイブ、66、355-365。
M.J. ミルゼンバーグ、A.R. レアード、S. セレン、C.S. カーター、D.C. グラーン (2009)。統合失調症における実行機能に関する 41 件の機能的神経画像研究のメタ分析。一般精神医学アーカイブ、66、811-822。
モンタギュー、R. (2006)。なぜこの本を選んだのでしょうか?私たちがどのように意思決定を行うか。ニューヨーク: Dutton Press、ネメロフ、CB (2004)。幼少期のトラウマの神経生物学的影響。臨床精神医学ジャーナル、65(補足 1)、18-28。
E.J. ネスラー (2005)。依存症に共通する分子経路はあるのでしょうか? Nature Neuroscience、8、1445-1449。
ニコルズ、D.E. (2004)。幻覚剤。薬理学と治療学、101、131-181。 Ongur, D.、Drevets, W.C.、および Price, J.L. (1998)。気分障害における前頭前皮質下皮質のグリアの減少。米国科学アカデミー紀要(米国)、95、13290-13295。
パンクセップ、J. (2006)。進化精神医学における感情エンドフェノタイプ。神経精神薬理学と生物学的精神医学の進歩、30、774-784。
ペソア、L.、アドルフス、R. (2010)。感情処理と扁桃体: 生物学的重要性の評価の「低い道」から「多い道」へ。 Nature Reviews Neuroscience、11、773-782。パイン、D.S.、ヘルフィンスタイン、S.M.、バーハイム、Y.、ネルソン、E.、およびフォックス、NA (2009)。小児不安障害の新しい治療法開発における課題: 不安障害に関する研究からの教訓。神経精神薬理学、34、213-228。
ピザガリ、DA (2011)。うつ病における前帯状回機能不全:治療反応のバイオマーカーに向けて。神経精神薬理学レビュー、36、183-206。
J.L. プライス、W.C. ドレベッツ (2010)。気分障害の神経回路。神経精神薬理学レビュー、35、192-216。
Ragland, J.D.、Laird, A.R.、Ranganath, C.、Blumenfeld, R.S.、Gonzales, S.M.、および Glahn, D.C. (2009)。統合失調症におけるエピソード記憶中の前頭前野活性化の欠陥。アメリカ精神医学ジャーナル、166、863-874。
ラマチャンドラン、VS.、ブレークスリー、S. (1998)。脳の中のファントム:人間の心の謎を探る。ニューヨーク:ウィリアム・モロー。
K.J. レスラー、H.S. メイバーグ (2007)。気分障害や不安障害における異常な神経回路を標的とする: 研究室から臨床へ。 Nature Neuroscience、10、1116-1124。 Rotge, J. Y.、Aouizerate, B.、Tignol, J.、Bioulac, B.、Burbaud, P.、および Guehl, D. (2010)。強迫性障害におけるグルタミン酸ベースの遺伝的免疫仮説。遺伝子から症状までの統合的アプローチ。神経科学、165、408-417。
Sambataro, F.、Blasi, G.、Fazio, L.、Caforio, G.、Taurisano, P.、Romano, R.、他(2010年)。オランザピンによる治療は、統合失調症患者におけるデフォルトモードネットワークの調節と関連しています。神経精神薬理学、35、904-912。
Satterthwaite, T.D.、Wolf, D.H.、Loughead, J.、Ruparel, K.、Valdez, J.N.、Siegel, S.J. 他(2010年)。統合失調症における脅威に対する大脳辺縁系反応の亢進と皮質顔認識記憶反応の低下との関連性。アメリカ精神医学ジャーナル、167、418-426。 J. サビッツ、W.C. ドレベッツ (2009)。双極性障害および大うつ病性障害: 発達と変性の分裂を神経画像化する。神経科学と生物行動のレビュー、33、699-771。
Schwabe, L.、Wolf, O.T. (2009)。ストレスは人間の習慣的な行動を促します。神経科学ジャーナル、29、7191-7198。
Seeley, W. W.、Crawford, R. K.、Zhou, J.、Miller, B. L.、Greicius, M. D. (2009)。神経変性疾患は、大規模な人間の脳ネットワークを標的としています。ニューロン、62、42-52。
Seeley、W.W.、Menon、V.、Schatzberg、A.F.、Keller、J.、Glover、G.H.、Kenna、H.、他。 (2007)。顕著性の処理と実行制御のための、分離可能な固有の接続ネットワーク。神経科学ジャーナル、27、2349-2356。
Seminowicz, D.A.、Mayberg, H.S.、McIntosh, A.R.、Goldapple, K.、Kennedy, S.、Segal, Z.、他(2004)。大うつ病における辺縁前頭回路: 経路モデリングメタアナリシス。ニューロイメージ、22、409-418。
Sheline, Y.I.、Barch, D.M.、Price, J.L.、Rundle, M.M.、Vaishnavi, S.N.、Snyder, A.Z. 他(2009年)。うつ病におけるデフォルトモードのネットワークと自己言及プロセス。米国科学アカデミー紀要(米国)、106、1942-1947。
Sheline, Y.I.、Price, J.L.、Yan, Z.、Mintun, M.A. (2010)。うつ病における安静状態の機能的MRIは、背側結合を介したネットワーク間の接続の増加を明らかにします。米国科学アカデミー紀要(米国)、107、11020-11025。
Shin、L.M.、および Liberzon, I. (2010)。恐怖、ストレス、不安障害の神経回路。神経精神薬理学レビュー、35、169-191。
Van den Heuvel, M.P.、Stam, C.J.、Kahn, R.S.、および Hulshoff Pol, H.E. (2009)。機能的な脳ネットワークと知的パフォーマンスの効率。神経科学ジャーナル、29、7619-7624。
Weiss、A.P.、Ellis、C.B.、Roffman、J.L.、Stufflebeam、S.、Hamalainen、M.S.、Duff、M.、他。 (2009年)。統合失調症における認識記憶中の前頭頭頂機能の異常: マルチモーダル神経画像研究。神経科学ジャーナル、29、11347-11359。
Whitfield-Gabrieli, S.、Thermenos, H. W.、Milanovic, S.、Tsuang, M. T.、Faraone, S. V.、McCarley, R. W.、他(2009年)。統合失調症および統合失調症患者の第一親等におけるデフォルトネットワークの活動亢進および過剰接続。米国科学アカデミー紀要(米国)、106、1279-1284。
Zanelli, J.、Reichenberg, A.、Morgan, K.、Fearon, P.、Kravariti, E.、Dazzan, P.、他。 (2010年)。特異的および一般的な神経心理学的欠損: さまざまな精神病の初発症状を呈する患者の比較。アメリカ精神医学ジャーナル、167、78-85。
