
4 脳ネットワークと人間の心
内因性接続ネットワーク (ICN) に関する背景情報を踏まえて、脳システムがどのようにして心を生み出すと考えられているのか、そして脳ネットワーク内および脳ネットワーク全体の機能不全がどのように精神症状や精神障害に寄与するのかを考えていきます。この議論では、前の章で概説され、ジョゼフ・ルドゥーが著書『シナプス・セルフ』で擁護した「心」の定義を引き続き使用します。ルドゥー氏によれば、心は脳回路の活動の結果であり、人間が考えること(認知)、物事に価値を付ける(感情)、目標を設定して達成する(動機)という3つのことを可能にします。すべての主要な精神疾患には、心のこれら 3 つの側面のそれぞれに欠陥が含まれているが、心の特定の側面の機能不全は、必ずしもそのシステムに局在する特定の病理があることを意味するわけではない、というのが私たちの前提です。むしろ、相互リンクされたシステムにおける効果的な処理は、他のネットワークからの適切かつ正確な入力に依存しており、あるシステムの病状が別のシステムの機能不全を引き起こす可能性があります。前に引用した 2 つの例がこのことを強調しています。まず、片麻痺などの切断症候群では、損傷したシステムからの適切な入力が欠如しているため、損傷を受けていない残りのシステムによる独特の解釈が生じます。健康な脳領域は、他の領域からの問題のある入力に直面したときに独特の解釈をする可能性があるというこの概念は、脳の働きの根底にある基本原則であると思われます。第二に、アルツハイマー病のような病気は、初期の特定のICNに対する優先的な攻撃によって最初に生じる可能性がありますが、臨床症状は、非常に初期の時点であっても、認知、感情、および動機におけるより広範な欠陥を伴います。これら後者の概念は、最近、局所病変による損傷を受けた脳ネットワークの接続性を調べる研究で拡張されています。単一のネットワーク (認知制御ネットワークなど) に限定された損傷は、損傷したシステム内の損傷を受けていないノードの機能不全や接続の変化を引き起こしますが、密接に関連しているが損傷を受けていないネットワーク内では再配置には至らないようです。それにもかかわらず、損傷したシステム内の処理に欠陥があるため、2 つのネットワーク間の相互作用は変化します。
認知(思考)
作業記憶と前頭前野
人間の認知力が非常に強力であることにはほとんど疑いの余地がありません。これにより、外界からの異なる入力を処理し、現在の健康状態や記憶の評価など、これらの入力を個人的な内界と組み合わせる能力が得られます。このような統合には、高度に接続された脳の領域、ルドゥーの用語では「収束ゾーン」の調整された活動が必要です。私たちが何を考えているかに気づくためには、その考えを「オンライン」で意識的に保持する必要があります。したがって、脳にこのプロセス専用のネットワークがあることは驚くべきことではありません。認知科学者はこれを「作業記憶」と呼んでいます。作業記憶は単一の脳領域の活動から生じるものではありませんが、前頭前皮質 (PFC)、特に背外側 PFC (dIPFC) が重要な役割を果たしています。私たちが意識的に何かについて考えているとき (今日の昼食に何を食べたのか、今夜何をしなければならないのかなど)、dIPFC はこの情報を扱っています。
ワーキングメモリは、意識的に考えるということの最も基本的な側面を表していると考えられます。これにはいくつかの重要な機能があります。まず、作業記憶は記憶装置ではありません。むしろ、複数のソースから情報を取得し、使用中にその情報を追跡する一連の操作であるように見えます。一種の「スクラッチ パッド」処理です。第二に、作業記憶の容量には限界があり、人が現在の考えを意識し続けてタスクを順調に進めるには、作業記憶を継続的に更新する必要があります。作業記憶は一度に約 5 ~ 9 個の項目を保持できると推定されています。心理学では、これは「7 プラスまたはマイナス 2」(7 ± 2) ルールと呼ばれ、これが 7 桁の電話番号の使用と、人間がそれほど効率的ではないという事実の認知的基盤であると考えられています。ほとんどのタスクには複数のサブコンポーネントがあるため、複数のタスクを同時に処理する場合があります。最近の研究によると、人間は自分はマルチタスクが得意だと信じていても、頻繁にマルチタスクを行う人、特に複数の種類のメディアを使用する人は、実際には非常に気が散りやすく、少ないことをやろうとする人よりもタスクを完了するのが難しいことがわかっています。特定の時間に、より限られたメディアのレパートリーを使用します。この気が散りやすい原因の 1 つは、作業記憶の過負荷と、PFC が自然に機能しているように見える方法を反映しています。作業記憶の容量は限られていますが、頻繁に更新される可能性があり、この更新は「考える」ときに思考の流れを維持する能力にとって重要である可能性があります。作業記憶がどのように継続的に更新されるかは、積極的な研究の問題です。いくつかの証拠は、NMDA 型グルタミン酸受容体に作用する入力の重要性と、フィルター装置としてのドーパミン D1 受容体の関与を示唆しています。 dIPFC における持続的な神経活動も寄与している可能性があります。
脳はどのように思考内容を選択するのでしょうか?
ワーキングメモリに関与するネットワークは意識的な思考にとって重要ですが、脳は思考の内容を選択する方法、つまり思考に何を含めるか、何を除外するかをどのように選択するかを理解する必要があります。このプロセスでは、少なくとも 2 つの他の ICN、デフォルト モードの ICN と注意を制御する ICN (または ICNS) が主要な役割を果たしているようです。作業記憶は注意ネットワークと密接に関連しており、この関連性を利用する少なくとも 2 つの重要な操作が関与しているようです。1 つは、吻側の上前頭溝、後帯状皮質、および楔前部に作用する項目を「選択する」メカニズムです。尾側の上前頭溝と後頭頂皮質を含む「更新」操作。この後者のシステムは、私たちの注意の焦点を変えるのに役立ちます(主要な脳領域の位置については付録を参照)。
第 3 章で説明したように、機能画像研究は、人間の脳が実際にはアイドル状態や休息状態にあることは決してないことを示しています。私たちが活動的な作業に従事していないときは、脳の特定の領域が代謝と血流の点で非常に活発になります。これが脳が非常に多くのエネルギーを消費する主な理由の 1 つです。タスク依存のエネルギー使用量は、このベースライン活動と比較して、全体の脳エネルギー需要に対する比較的小さな寄与 (5% 以下) です。人の注意が関心のあるタスクに向けられると、ベースラインでの使用量が多い領域の活動が減少し、特定のタスクに必要な領域の脳活動が増加します。特定のタスクに必要な集中した注意から、より「リラックスした」脳状態に移行すると、そのタスクに必要な脳領域の活動が減少し、バックグラウンド活動が再び増加します。人間がさまざまな種類の情報をどのように処理するかを調べるさまざまな機能画像研究により、同じ一連の脳領域がこの高いベースライン (バックグラウンド) 活動に関与していることが判明しました。第 2 章と第 3 章で述べたように、これにより Marc Raichle らはこの脳ネットワークを「デフォルト システム」と呼ぶようになりました。デフォルトモードネットワークには、腹内側 PFC、後帯状皮質および後脾皮質、楔前部および腹内側側頭葉の一部、海馬、および下頭頂小葉を含む、接続された脳領域の分散集合が含まれます。図2-1)。重要なのは、これらの同じデフォルト領域の一部が作業記憶システムと重複していることです。私たちが起きているときは、特定のタスクを実行していないときでも、通常何かについて考えていることを考えると、これは驚くべきことではありません。
私たちがタスク(数学の問題を解く、この文章を読むなど)に注意を集中するときは、作業記憶を活用し、デフォルトの状態から、目の前の仕事を実行するために必要な ICN を活用するモードに移行する必要があります。この注意を移すプロセスの重要なコンポーネントは、マウリツィオ コルベッタが「方向転換システム」と呼ぶ ICN です。視覚系では、これには、前頭葉と頭頂葉を結び付ける右 (非優位) 半球の皮質回路が含まれます。それは少なくとも 2 つの経路で構成されます。1 つは現在の思考を中断し、新しいタスクへの注意をリセットする腹側回路、もう 1 つは新しい注意対象を選択し、必要な計算に適切な ICN とそれらをリンクする背側経路です。図 4-1 は、背側と腹側の再配向ネットワークの図を示しています。
腹道は、私たちの認知の焦点を変え、注意を移すことを可能にする一種の「サーキットブレーカー」のように機能します。これはタスクに関連した刺激によってトリガーされ、デフォルトのシステムの内部焦点から特定の外部要求に対処する操作に切り替えることができます。背側経路は、注目すべき特定の項目を選択し、目の前の特定の刺激に関連する作業記憶の内容を決定するために外部変数の性質を評価するのに役立ちます。言い換えれば、背側システムは外部(世界)の要求を優先するのに対し、デフォルトシステムは内部(自己)の情報を優先するようです。おそらく、作業記憶の容量が限られているため、通常、私たちはこれらのタスクの両方を一度に実行することはありません。
————————図 4-1
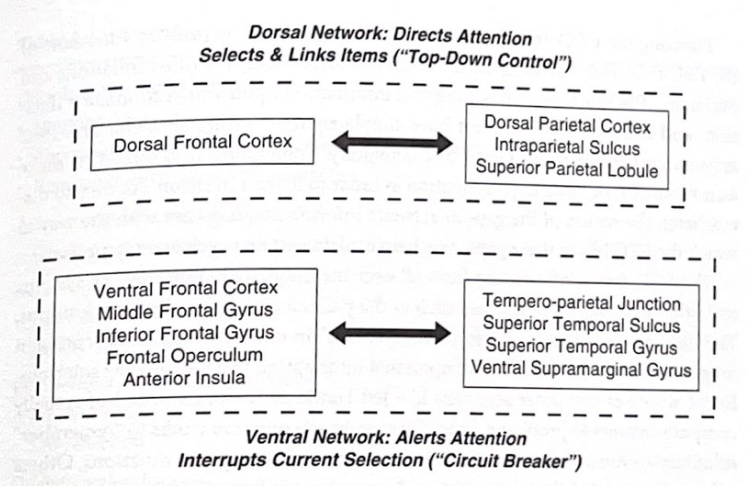
背側ネットワーク: 注意を向けてアイテムを選択およびリンクします (「トップダウン制御」)
背側頭頂皮質 頭頂内溝 上頭頂小葉
背側前頭皮質
腹側前頭皮質 中部前頭回 下前頭回 前頭蓋蓋 前島
側頭頭頂接合部 上側頭溝 上側頭回
腹縁上回
腹部ネットワーク: 注意を喚起する
現在の選択を中断します (「サーキットブレーカー」)
図 4-1 注意と前頭頭頂部の方向転換ネットワーク。この図は、Maurizio Corbetta らによって説明された方向転換と注意ネットワークにおいて重要であると考えられる主要な脳構造を示しています。
————————-
同時に、あるいは少なくとも両方を同時に効率的に実行することはできません。言い換えれば、空想は通常、仕事を成し遂げるのに役立ちません。
PFC はワーキングメモリ以上の役割を果たします
PFC は、作業記憶と注意における役割に加えて、計画と意思決定、推論の引き出し、因果関係の決定、物、人、および物事についての予測を含む、他の多くの認知タスクにも重要です。規則性とパターンを判断することによってイベントを分析します。これらは脳のいわゆる「実行」機能の一部であり、認知処理の最高位レベルを表します。
この実行機能のアナロジーを使用して、エルコノン ゴールドバーグは PFC を脳の「最高経営責任者」(CEO) と表現しました。企業の CEO に当てはまることですが、このアナロジーは、PFC がおそらくすべてのジョブ (計算) を実行するサイトではなく、特定の問題を解決するために必要なスキルを備えたサイトでさえないことを示しています。むしろ、PFC は、組織内の「誰」が特定のタスクに必要な能力を持っているかを認識し、処理のためにその「ジョブ」をそれらの領域に参照します。結果はその後、さらなる分析、評価、意思決定のために PFC にフィードバックされます。 PFC のさまざまな部分は、脳収束ゾーンの中で最も高度に接続された領域の 1 つであり、そこでは多くのソースからの入力が結合、統合され、応答を生成するために他の脳領域に信号を送るために使用されます。人類の進化の過程における PFC のサイズと接続性の大幅な拡大は、他の種と比較してヒトが示す高度な認知能力の主な要因の 1 つであると考えられています。
CEO の例えをさらに追求すると、PFC の主な機能の 1 つに、ほとんどの状況や意思決定に伴う不確実性への対処が含まれると考える人もいます。脳が意思決定を下すための完全な情報を持っていることはほとんどなく、ほとんどの問題には単純な正解または不正解はありません。 PFC は、この曖昧さに対処するのに適していると思われます。場合によっては、意思決定を行うために、不足している情報について推測を行わざるを得なくなることがあります。後で説明するように、正確な情報のギャップの性質と、PFC がこれらのギャップを埋める方法は、特定の精神症状を説明するのに役立つ可能性があります。
PFC は、感覚系や、頭頂皮質や海馬などの他の主要な収束ゾーンを含む、脳全体から情報を引き出します。 PFC は、新皮質の他の領域の長期記憶に「保存」されている情報にもアクセスでき、その情報を呼び出して新しい解決策を生成することができます。ジェフ・ホーキンスのようなコンピューター科学者の言葉を借りれば、脳は実際に問題に対する答えを計算するわけではありません。むしろ、解決策を「記憶」するために、さらには新しく曖昧な状況に対する解決策を「記憶」するために、記憶バンクを検索します。これを PFC の「未来を記憶する」能力と呼ぶ人もいます。未来は私たちの保存された記憶にとって実際には何の意味もありませんが、PFC はしばしば海馬と連携して機能し、以前の情報を利用して新しい状況に適用しようとします。
PFC のさまざまな機能により、この脳領域は内部の解剖学的構造とその入出力の点で非常に複雑になっていることが想像できます。詳細に迷わないように、PFC を外側、内側、腹側の 3 つの主要なサブセクションに分割する単純なスキームを使用します。側方 PFC、特に背外側面は作業記憶に大きく寄与しているようです。それは高等霊長類と人間で最も発達しています。内側 PFC には前帯状皮質が含まれており、意思決定と実行する出力の選択に関与します。これは、脳の真の「実行」領域である可能性があります。腹側(眼窩)PFC は脳の感情処理システムと結合しています。それは感情が意思決定に影響を与える方法を提供し、ひいては感情を制御するのに役立ちます。 PFC は脳全体に多様な遠心性 (流出) 接続を持ち、複雑な一連の反応の生成を可能にします。表 4-1 と図 4-2 は、PFC 内のサブ領域とその提案される認知機能の説明を示しています。
重要な PFC 接続の 1 つは、優位な (通常は左) 大脳半球の言語に特化した ICNS との密接な結合です。この洗練された言語 ICNS との結合は人間で最も高度に発達しており、人間に言語の観点から考える能力を与えます。言語は、人間が単純な認識を超えて複雑な概念の領域に移行できるようにする、概念的な比喩、類推、カテゴリータイプの思考を生成するための基礎を提供します。興味深いことに、これらの高次の抽象概念の多くは、より単純な感覚システムの動作と結びついており、それによって私たちは抽象的な概念 (愛情や愛など) を感覚用語 (暖かい、熱い、冷たい) で表現することになります。このタイプの抽象化は私たちにとって非常に自然なため、ほとんどが当然のこととして受け入れられており、数学や科学の概念 (たとえば、数値は直線上の点や位置の値であるという考え) を含む、私たちの抽象的な概念の多くに浸透しています。ゲーム内)。人間の思考を理解するには、多くの場合、概念を表現するために使用されている比喩を解読する必要があります。一部の精神疾患では、このタイプの抽象化の欠陥により、「具体的な」(硬直した)思考やコミュニケーションの問題が生じる可能性があります。精神科医は、患者に簡単なたとえやことわざを解釈してもらうことで、これを評価しようとすることがあります。比喩や抽象化を誤解すると、その人が伝えようとしているメッセージをまったく理解できなくなる可能性があります。言語 ICN との対話を介して応答を生成することに加えて、PFC は他の領域に応答を指示する機能を備えています。これは一般に「トップダウン処理」と呼ばれ、この制御モードが感情、動機、運動機能などのより原始的なシステムを制御するために脳の最高レベルを使用するという事実を反映しています。 PFC がトップダウン制御を発揮するために、PFC を介した認知制御を達成するために、異なる時間スケールを利用するいくつかの ICN が進化しました。このような ICN の 1 つは、焦点を迅速に変更し、制御を調整する手段を提供する前頭頭頂系に関係します (注意を伴う動作と同様です)。 2 番目の ICN には、PFC の鰓蓋領域と帯状皮質が関与します。この帯状蓋システムは、作業期間中安定した維持を提供することにより、より永続的な制御の役割を果たします。 (前頭の「鰓蓋」は、下前頭回の最も後方の部分であり、優位半球のブローカ言語野を含みます。)これらのシステムはどちらも、作業記憶や他の領域に関与する PFC の構造を利用しています。意思決定に関与します。主に運動機能に関与する構造である小脳が、前頭頭頂制御システムと帯状鞋蓋制御システムの間の相互作用の仲介に関与している可能性があるという証拠もある。この後者の観察は、神経回路がいかに複雑であるか、そして脳の領域を「運動」、「感覚」、または「認知」として定義することがいかに恣意的で機能に依存する可能性があるかを示しています。
————————-
表 4-1 前頭葉: 細分化と提案された機能
背外側前頭前野 (dIPFC)
・ワーキングメモリ
・注意、感情、衝動をトップダウンでコントロールする
・推論と曖昧さへの対処
前帯状皮質 (ACC)
・競合監視
背内側前頭前野 (dmPFC)
・エラー検出
・リアリティモニタリング
・腹内側前頭前野(vmPFC)
・感情の調整
・反省
眼窩前頭皮質 (OFC)
・刺激の感情値と報酬の期待値
・インパルス制御
————————図 4-2
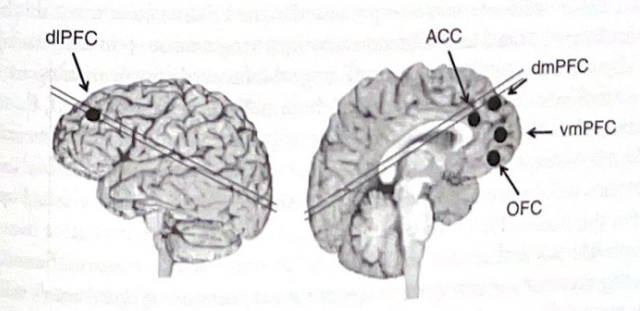
図 4-2 前頭葉の主要な領域。この図は、表 4-1 にリストされている PFC の主要な領域のおおよその位置を示しています。略語については、表 4-1 を参照してください。 (許可を得て、Damasio、2005 年から翻案。)
————————-
これら 2 つの認知制御システムは、機能を調節するために多くの戦略を利用します。 2 つの例は、「プロアクティブ」戦略と「リアクティブ」戦略と呼ばれます。積極的な戦略は、注意、認識、行動を目標に向けて偏らせますが、事後的な戦略は、一種の誤り訂正である変更の必要性を検出した場合にのみ反応します。興味深いことに、年齢は認知制御に使用される戦略に影響を与える可能性があり、高齢者はより反応的であり、若い人はより積極的です。また、個人が自分の好みのコントロール方法に適応して変更するように教えることもできるようで、これは心理療法やリハビリテーション戦略のターゲットとなる可能性がある。感情とモチベーションをトップダウンでコントロールするのは非常に難しく、エネルギーを必要とする可能性があることを強調することが重要です。より原始的な脳の中枢は、この種の制御を行うように設計されておらず、それらが PFC とは独立して反応や行動を生成できることは明らかです。 LeDoux 氏が指摘したように、これが、正しいことを知り、実行することが難しい理由である可能性があります。図 4-3 は、いくつかの形式のトップダウン処理がどのように動作すると考えられるかを強調した図を示しています。
PFC および精神神経障害
複雑な高次情報は PFC で処理されるため、この脳領域は精神神経疾患に伴う機能障害に関与することがよくあります。 PFC 機能の異常 (意思決定や感情のトップダウン制御など) は PFC 自体の異常を示唆している可能性がありますが、必ずしもそうとは限りません。むしろ、PFC は、他の領域から誤った情報を受け取ったり、主要な入出力から切断されたりするために、パフォーマンスが低下する可能性があります。 PFC への入力または出力に問題があると、臨床症状が発生する可能性があります。このため、エルコノン・ゴールドバーグ氏は、実行機能の欠陥を「発熱」などの非特異的な身体症状に例えています。言い換えれば、異常な実行処理は、脳機能に何か問題があることを示しています。ただ、何が問題なのか、主な問題がどこにあるのかはわかりません。
曖昧さに対処することの困難は、精神疾患における PFC 機能不全の初期の症状の 1 つである可能性があります。たとえば、あいまいな情報や混乱を招く情報は、奇妙に見える行動につながる可能性があります。不確実性への対処に問題があると、PFC が下位の脳領域に、そのタスクに不適切な出力で応答するように指示する可能性があります。たとえば、PFC は、ある人の靴がすべて紛失しているという誤った情報を受け取る可能性があります。その反応は、家族または未知の侵入者が靴を盗んだという告発である可能性があります。実際には、その人は記憶障害があり、靴をベッドの下に隠したことを忘れていました。出力には妄想的な性質がありますが、おそらく PFC が正しい結論に達するために必要なデータの一部のみを受信したという事実に起因するものと考えられます。同様に、PFC は、不確実でストレスを引き起こす状況に対処するために、定型的な行動反応 (例: 癇癪の爆発) を指示する場合があります。 「ストレスを感じている」人は、新たな課題に直面したときに、高次の目標に向けた行動よりも、習慣的な(暗黙的に学習された)反応を使用する傾向があります。これは、習慣的な反応には事前の検討や計画がほとんど必要ないという点で、エネルギーを節約する戦略です。それにもかかわらず、これらの応答は多くの設定において不適切である可能性があります。これは、特定の種類のパーソナリティ障害を患っている人に特有の問題である可能性があります。特に、その人が次のような傾向にある場合、過剰に学習した常同的な反応が日常生活を支配しているように見えます。
強調した。
————————-図 4-3
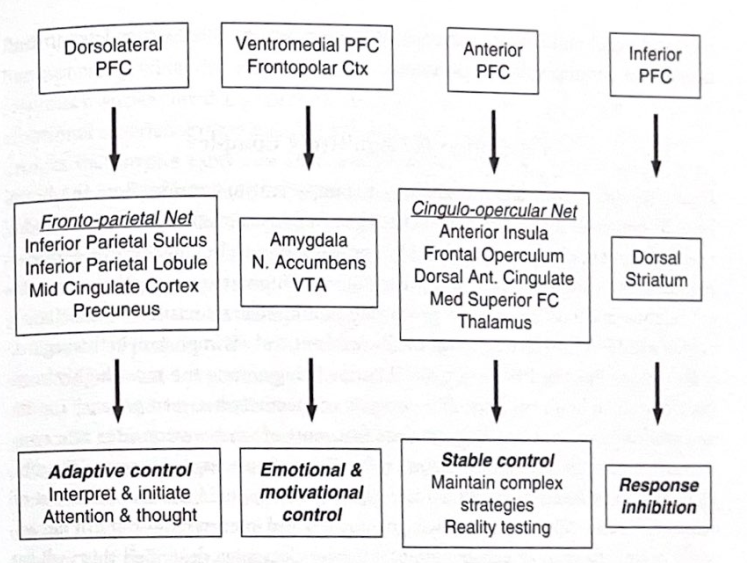
背外側PFC
前頭頂頂網 下頭頂溝 下頭頂小葉 中部帯状皮質 楔前部
適応制御 解釈と開始 注意と思考
腹内側 PFC 前極 Ctx
前部PFC
劣る
PFC
扁桃体
No. リクライニング VTA
ベルトカバーネット前部アイランド
前頭蓋背側アリ。シングレート メッド スーペリア FC 視床
背側線条体
感情とモチベーションのコントロール
安定した制御 複雑な戦略の維持 リアリティテスト
応答阻害
図 4-3 トップダウン処理: 前頭前野と制御ネットワーク。この図は、精神処理をトップダウンで制御する主要な PFC 領域とネットワークの一部を示しています。本書で説明され、Steve Petersen らによって説明された適応制御ネットワークと安定セット制御ネットワークが含まれています。キー構造の場所については、付録を参照してください。
————————-
PFC 機能の欠陥を検出する際の大きな問題の 1 つは、PFC の高次処理に関する現在の臨床検査が、運動機能、感覚機能、または言語機能の評価に比べて比較的粗雑であることです。臨床検査やより高度な神経心理学的検査は、現実世界のあいまいな状況で患者がどのように反応し、意思決定を行うかを判断するよりも、作業記憶や注意力の欠陥を監視することに優れています。これらの問題については、後の精神疾患に関する議論で再び取り上げます。
知覚は認知的に複雑です
人間の認知について議論する際には、脳が実際に入力をどのように認識するかを考慮することが重要です。脳は、単峰性タイプの入力(たとえば、個々の一次感覚からの入力)を処理する洗練されたシステムを備えていますが、より高次の処理には、一次視床皮質領域への単峰性入力と、多峰性の関連(連合皮質と連合皮質)を処理する皮質領域との間の複雑な相互作用が含まれます。 PFC)。この点では、スモールワールドネットワークの構造が重要であり、PFCや頭頂皮質などの領域は、脳回路の中で最も高度に接続されている領域の1つです。一次入力と作業記憶の間のこの複雑な接続により、私たちが意識的な「認識」とみなしているもののほとんどが、高度に処理された情報であることがほぼ保証されます。一部の認知科学者は、実際には私たちの脳には一次的な知覚はほとんどなく、意識に入ってくるもののほとんどは高度にフィルタリングされ解釈されているが、事実の観点から必ずしも正しいわけではないと結論付けています。マイケル・ガザニガは、脳を「自己関心のある通訳者」と呼んでこれをうまく説明しました。私たちが私たちの周りで、そして私たちに起こっていると信じていることは、実際には、感覚入力、記憶、コンテキスト、内部状態 (感情と動機) の混合物であり、受信データを分類して単純化するための抽象化技術を使用して PFC によって解釈されます。後者の例には、新しい経験についての考えを伝え、整理するのに役立つ抽象的なカテゴリー (「植物」、「動物」、または「スポーツ」など)、比喩、類推の使用が含まれます。実際、高次処理には高価なエネルギーが必要となるため、脳、特に PFC は近道を選択し、その記憶を使って経験していることを予測する必要があります。脳は一度に全体像を捉えることはほとんどなく、むしろシーンの一部に焦点を当て、残りについて「推測」します。グレゴリー・バーンズ氏が指摘したように、これらのショートカットは脳が過負荷を回避する方法を表しています。このタイプの処理を使用すると、脳は間違いを犯したことに気づいた場合にのみ認識を変更します。間違いを認識し、「エラー信号」を生成することは、知覚のプロセスの非常に重要な部分であり、エラー修正に欠陥がある限り、誤った認識が意識経験を支配する可能性があります(機嫌の悪さ、妄想など)。内部エラー修正は認知的な「現実チェック」の一種であり、複雑な状況に対処するための基本です。
側方化された脳機能と認知
脳は異なる処理モードを使用して世界についての予測的解釈を行い、2 つの大脳半球は相補的だが異なる方法で情報を処理しているようです。この概念は、さまざまな機能が皮質でどのように左右化されるかを考えるときに重要です。これは、左(優位)半球で処理される言語機能に関して最も明確です。対照的に、空間情報は右 (非優位) 半球で優先的に処理されます。
感情は新皮質でも異なって処理されるようで、右半球が感情反応の解釈と生成に主要な役割を果たしています。右半球が関与する脳卒中は、感情経験の欠陥と関連しており、その結果、数種類の失語症が引き起こされる可能性があります。失語症は、感情信号伝達における表現および/または受容の問題を伴う臨床症候群です。欠陥の例には、特定の感情を表現できないこと(例、悲しいというよりも当たり障りのない、または鈍感に見える)や、他人の感情を解釈するのが難しい(怒りを別の感情状態と間違えたり、感情が表現されていることさえ認識できない)などが含まれます。どちらのタイプの問題も、社会的コミュニケーションや対人関係に大きな欠陥を引き起こす可能性があります。
「分裂脳」患者の研究は、大脳半球における側性処理を理解する上で非常に有益である。通常、これらの人は難治性てんかんを患っており、発作が半球から半球に広がるのを防ぐために脳梁を外科的に切断されています。脳梁は、脳の中央にある大きな線維束で、2 つの大脳半球を接続しています。分割脳個体の研究により、半球がどのように孤立して機能するかについて独自の洞察が得られました。左半球は、その反応に論理性と一貫性を求めるようです。事実であることや正確であることには関心がなく、一貫したストーリーを伝えることだけに関心があるようです。通常、自分が何かを知らないことを認めず、必要に応じて答えを「でっち上げ」ます。たとえば、右半球の損傷に起因する片麻痺切断症候群では、損傷を受けていない左半球によって、麻痺した手が別の誰かのものであるというストーリーが生成されます。対照的に、右半球は正確な詳細に関心があり、矛盾に対処するのが難しいようです。残念ながら、この半球には直接的な言語モジュールがないため、感情や気持ちを通じて表現する傾向があります。両半球間のこうした違いは、前部左半球に関わる脳卒中が右半球の病変よりもうつ病とより一般的に関連しているという観察に寄与している可能性がある。左側が損傷すると、右半球は問題があることを認識し、「心配」になり、否定的な感情を通じてその懸念を表現します。右半球に同様の位置に前方病変がある場合、左半球は通常問題なく業務を遂行できますが、右半球によって課せられた感情制御によって抑制されません。これにより、一部の人では躁状態のような(または衝動的な)行動が発生する可能性があります。
ケース。
特に論理と感情に関して、なぜこのような半球機能の違いが進化したのかは明らかではありませんが、部分的には、自律神経系からの情報が脳内で処理される方法を反映している可能性があります。覚醒と生存に関するデータを伝える交感神経系からの入力は右前脳で優先的に処理され、リラックスと親近感を伴う副交感神経系からの入力は左前脳で処理されます。興味深いことに、これらの異なる入力と処理モードには異なるエネルギー要件があるようで、左側 (全体的) の処理はよりエネルギーを節約し、右側 (詳細指向) の処理はよりエネルギーを大量に消費します。
知性と認知の柔軟性
認知に関するこの最初の議論を終える前に、人間の知性とその精神障害との潜在的な関連性についていくつかコメントしたいと思います。 「インテリジェンス」とは、正確に定義するのが難しく、政治的な意味を持つ概念です。これはさまざまな脳領域と ICN の寄与を反映しており、脳の新たな特性のようなものです。議論はありますが、さまざまなタイプの知性が存在する可能性があり、これらの領域では個人が大幅に異なる可能性もあるようです。これらには、言語、数学と論理、音楽、運動/運動、空間関係、社会的機能に関わる能力が含まれます。社会的知性は、自分自身の心と他人の心を理解する能力を反映しています。感情的理解と共感は、一般的な社会的知性のサブセットである可能性がありますが、別の形態の知性を構成する可能性があります。これらのさまざまな形の知能は測定が難しい場合があり、言語的および非言語的知能指数 (IQ) に基づいて臨床的に使用されている多くの手段は文化的な偏見にさらされており、多くの点で望ましくない点が残されています。それにもかかわらず、IQ パフォーマンスの欠陥は、教育、社会、職業活動を妨げる可能性のある機能障害の領域を特定するのに重要な場合があります。また、IQ テストの成績が高いからといって神経精神疾患を予防できるわけではありませんが、知的能力が低いと、うつ病、薬物乱用、精神病などのいくつかの主要な疾患のリスク増加と関連しています。同様に、知的能力は、認知症疾患のリスクが最も高い人や認知症の経過を予測するために重要である可能性があります。この一例は修道女研究から来ています。これは、閉鎖的な修道会で暮らす女性の長期的な成果を調査した縦断的研究です。成人初期に、修道院に入る人々は自伝を書くように求められました。より高度で複雑な文体を書いた女性は、認知能力と認知症のリスクの点で、年齢を重ねてもより良い成績を収めた。この違いの理由は定かではありませんが、より高い知的能力が病気の緩和に役立つ可能性があるという「認知予備力」仮説と一致しています。トップダウンの処理と感情のコントロールも、より高い知的能力から恩恵を受けるようです。それにもかかわらず、ある領域で「賢い」からといって、他の領域での知性が必ずしも予測できるわけではありません。また、知性の形態が異なると、トップダウンで処理し、感情やモチベーションを制御する能力も異なるかどうかは不明です。また、「知性」の基礎となる脳回路には複数の認知プロセスが関与しており、これらの機能には前頭葉、頭頂葉、側頭葉の皮質における計算とこれらの領域間の相互接続が含まれることが多いことも明らかです。特に、高度なインテリジェンスは、前述したスモールワールド ネットワーク処理の利点を反映している可能性があります。
人間は本質的に社会的な動物であり、他者と関わる能力は人生の満足度に大きく関係しています。個人の社会的ネットワークやサポートシステムの欠陥は、精神障害の結果にも重大な影響を与える可能性があります。したがって、社会的知性の領域は精神医学において重要です。特に、他人が自分の心(経験と主体性)を持っていることを認識し、顔の表情、話し方、口調、非言語的な手がかりに基づいて他人の精神状態を推測する個人の能力は、他人と社会的につながる上で重要です。満足のいく対人関係を築く上で。この能力を理解することで、「心の理論」(TOM) という概念が生まれました。つまり、他の人が実際に何を考え、何を意味しているのかについての「理論」を構築します。 TOM は、人間はしばしば (常にではないにしても) 他人の心を「読もう」と試みていることを示唆しています。精神病患者にとって、そのような考えがどれほど煩わしいものであるかを考えると、これは興味深い概念です。精神病患者は、人に心を読まれたり、頭に考えを植え付けられたりするという考えにしばしば悩まされます。お互いの心を読むことは、私たちがどのように関わり、他者の意図、行動、感情、言葉を理解していくかについての基本的な信条です。内側および下側 PFC および上側頭溝(おそらく紡錘状回の顔面領域を含む)の活動は、TOM 処理に大きく寄与しているようです。興味深いことに、PFC の一部には、TOM タイプの処理に特に優れていると思われるニューロン (「ミラー ニューロン」と呼ばれる) があります。これらのニューロンは、他人の知覚された行動に反応して発火し、他人の動作(動きを含む)で知覚されたものを模倣する行動を開始します。 TOM の欠陥は複数の精神障害で観察されていますが、社会的愛着と互恵性の主要な欠陥が主要な特徴である自閉症で最も顕著であると考えられます。自閉症スペクトラム障害を持つ人は、他人の主体性(計画、意図、自制心)を認識する能力が低下しているようです。同様に、精神状態に関する誤った帰属は、他の障害における一種の認知欠陥を表している可能性があります。たとえば、無生物への主体性の帰属は、統合失調症的思考(「魔術的」思考と呼ばれる)で観察されます。
感情: 価値と意味の計算
どのような価値観、どのような意味があるのでしょうか?
ルドゥーの計画における心の 2 番目の要素である感情処理により、人間 (および他の動物) は遭遇したものに価値を付けることができます。感情に関与する脳システムは、PFC に比べて進化的に古いものです。げっ歯類でさえ、霊長類や人間に見られるネットワークと同様の方法で組織化された感情処理システムを持っています。これらのシステムが生存において重要な役割を果たしていることにほとんど疑いの余地はありません。感情によって、状況を迅速かつ無意識に評価し、それが安全か脅威かを判断することができます。これらは、私たちが「直感的な」決定を下すことを可能にする計算です。感情システムも、活性化されると脳の機能を制御し、行動を駆動するように設計されています。例えば、突然物音を聞いてびっくりして、何が起こっているのかを意識する前に防御姿勢を取ってしまうという経験は誰しもあるでしょう。最初の感覚入力 (視覚または聴覚) は皮質下で処理され、主要な生存システムを活性化します。皮質は二次的に関与し、運動行動に意識的な制限を加えることができます(認知用語では「反応阻害」と呼ばれます)。たとえば、意識的にそうしていることに気づく前に、私たちはヘビから遠ざかってしまうかもしれません。ヘビから離れた後、私たちは危険を理解し、より安全な場所に移動することを意識的に決定します。
人間には、幸福、悲しみ、恐怖、怒り、驚き、嫌悪という 6 つの「主要な」感情があるということである程度の合意はあります。一部の計画には軽蔑も含まれます。これらは 2 つの理由から一次感情とみなされます。1 つは、すべての人類集団で発生するように見えること、もう 1 つは、異なる文化の人々でも同様の方法で (同様の表情を使用して) 表現されることです。しかし、人間の感情生活は一次感情よりもはるかに豊かであり、社会的相互作用に関連する二次感情もたくさんあります。これらには、罪悪感、恥、当惑、嫉妬、誇り、愛などが含まれます。これらの二次感情は私たちの生活に大きな影響を与えますが、一次感情と同じように処理されるかどうか、また一次感情からどのように派生するかは明らかではありません。
感情。
原始的な脳システムが感情処理の根底にあることを考えると、他の動物の観点から感情について考えることも役に立ちます。 Jaak Panksepp は、動物には 7 つの主要な感情システムが存在することを説明しました。これらには、欲望(性的接近)、世話(母性の養育)、喜び(遊び)、恐怖(危険)、激怒(怒り)、パニック(別居苦痛)、および探求(探索)のネットワークが含まれます。人間の感情と同様に、パンクセップ氏は、これらのシステムは、遭遇が生命を維持するものであるか、生命を脅かすものであるかに関する情報を迅速にコード化し、適切な行動に対する適応的かつ本能的な反応を促進すると考えています。パンクセップはさらに、動物におけるこれらの主要なシステムは人間にも拡張されており、精神疾患の基礎を説明するための潜在的な「エンドフェノタイプ」として機能する可能性があると主張した。エンドフェノタイプは、複雑な病気の表現型(障害や症候群など)よりも遺伝子や神経系に密接に関係している可能性が高い、観察可能な(そして通常は定量化可能な)形質です。
感情はどのように処理されるのでしょうか?
すべての感情は、入ってくる情報の分析とその分析に基づくその後の出力を含む脳の計算を表します。このモデルでは、刺激は最初に意識的に (皮質で) または無意識的に (皮質下構造で) 知覚されます。特定の脳システムがこの知覚を処理し、身体反応を生成します。この反応は、多くの場合、基本的な身体生理機能 (心拍数、血圧、呼吸、体温など) を調節するシステムである自律神経系の出力の変化です。こうした身体の変化は脳によっても検出され、計算ミックスに組み込むことができます。たとえば、恐怖を感じると心拍数が増加します。これは脳によって認識され、正のフィードバック ループで心拍数がさらに増加します。身体感覚と文脈が作業記憶において意識的に認識される時点で、私たちはアントニオ・ダマシオの言う「感情」、つまり感情状態の意識的表現を経験します。この処理パスに沿ったいくつかの場所で、感情に応じて行動をとることができます。これは、私たちが「感情」を意識的に認識すると起こるかもしれません。その時点で、PFC は自発的な行動を促すことができます。あるいは、おそらくより興味深いことに、行動反応は、心拍数や呼吸の変化の前、または変化と同時に、新皮質が関与する前、そして何が起こっているのかを私たちが意識する前に発生する可能性があります。これは感情システムの主要な原則の 1 つです。つまり、感情システムは計算を行ったり、行動に変化をもたらすために皮質を必要としません。このため、これらは強力であると同時に、潜在的に問題を引き起こす可能性もあります。
これはどうして起こるのでしょうか?感情の処理に寄与する皮質下ネットワークと皮質ネットワークがいくつかあるようです。 LeDoux、Michael Davis、Michael Fanselow、その他多数の研究の結果、最も詳細に理解されているシステムは、げっ歯類でも同様に明確に経験される一次感情である恐怖の処理に関与するシステムです。人間。恐怖の処理では、側頭葉の奥深くに複数の亜核を持つ複雑なクルミ形の脳構造である扁桃体が主要な役割を果たします。恐怖を引き起こす刺激に遭遇すると、情報は最初に、感覚入力やその他の入力を処理するための皮質下の中継点である視床を経由して送られます。視床は、入力を迅速に評価するために扁桃体に直接接続し、さらに詳細な評価と解釈のために感覚新皮質にも入力を送信します。この単純な経路により、扁桃体は、皮質が関与する前に、視床下部および脳幹のストレスおよび警報システム、さらには線条体および運動系への自身の直接接続を介して防御行動を駆動することができます。シナプス接触とシナプス遅延の性質により、データを皮質に取得し、皮質内で作業記憶に分配するにはより多くのシナプスが必要であることを考慮すると、扁桃体が皮質よりも先に情報を取得することが保証されます。これに加えて、最近の研究では、扁桃体には独自の直接的な化学感知特性もあり、酸感知イオンチャネル(ASICS)を使用して脳の二酸化炭素レベルまたは酸塩基平衡の変化を検出していることが示されています。したがって、扁桃体は独自の直接的な感覚処理を行うことができます。その結果、恐怖を含む急速な感情反応が引き起こされる可能性があります。これが、呼吸の変化(過呼吸)が感情と密接に結びついている理由の 1 つである可能性があります。図 4-4 は、恐怖 (および不安) の処理に関与する神経回路と扁桃体の中心的な役割を強調しています。
視床 (または皮質) からの直接入力は、初期処理のために扁桃体の外側核に流れます。外側核は、感情処理を調節し、覚醒、防御行動、ホルモン反応を制御する領域への出力を生成する中核および側底扁桃体核に直接接続しており、中核は他の脳への出力のドライバーとして重要な役割を果たしています。地域。これらの出力領域には、覚醒と動機を支配する脳幹核(ノルアドレナリン、セロトニン、ドーパミン伝達物質系など)のほか、視床下部の神経内分泌ホルモン反応を調節する系や、水道周囲灰白質におけるすくみ反応(行動抑制)を媒介する系が含まれる。 。扁桃体は、視床下部の室傍核(PVN)との結合を介してコルチゾールなどのストレスホルモンの重要な活性化因子であり、コルチコトロフィン放出因子(CRF)の放出をもたらし、その結果、下垂体を刺激してACTHを放出します(副腎皮質刺激ホルモン)。 ACTHは副腎に作用してコルチゾールの分泌を促進します。コルチゾールは、フィードバック方式で脳に作用するなど、ストレス反応を仲介するために多くの働きをします。興味深いことに、コルチゾール分泌の調節の変化(日内変動の変化とデキサメタゾンによる抑制の減少)は、大うつ病やストレス関連障害において最も再現される生物学的所見の一つです。
————————-図 4-4
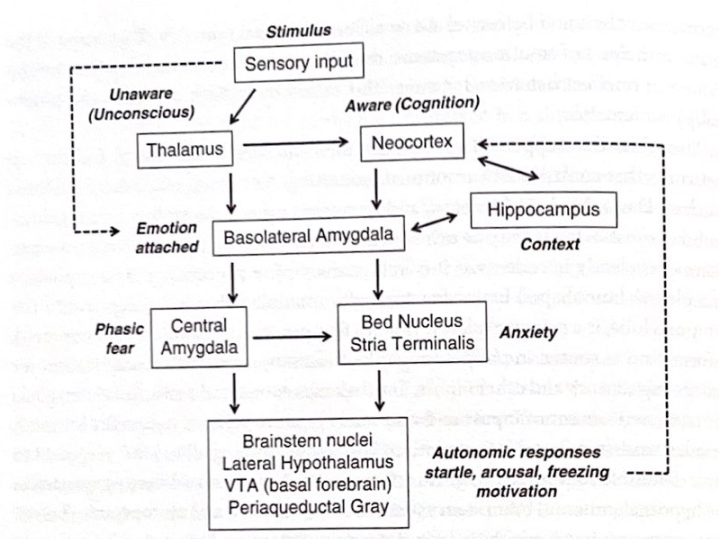
D
60 いいえ
刺激
無自覚(無意識)
感覚入力
意識(認知)大脳新皮質
視床
感情を伴う
扁桃体基底外側
海馬!
コンテクスト
一時的な恐怖
扁桃体中央部
ベッド核終端線条体
不安
脳幹核 外側視床下部 VTA (前脳基底部) 中水道周囲 灰色
自律反応は驚愕、興奮、モチベーションの低下を引き起こす
図 4-4 恐怖と不安。この図は、恐怖と不安の処理に関与する神経回路を示しており、扁桃体の中心的な役割を強調しています。この回路の多くは恐怖と不安の間で共有されていますが、不安に関与する回路は終端線の床核を含む拡張扁桃体に関与していると現在考えられています。キーの場所については付録を参照してください。
構造物。
————————-
扁桃体はシナプス可塑性も備えているため、恐怖を「学習」することができます。恐怖の刺激に繰り返しさらされると、無害な刺激が恐怖を生み出す刺激と関連付けられる一種の条件付けが生じる可能性があります。これはパブロフ条件付けの基礎であり、人間における同様の暗黙の効果が、特定の気分障害や不安障害に関与している可能性があります。扁桃体はまた、宣言的記憶の形成に重要であり、新規性や文脈の処理に役立つ脳領域である海馬との強力な双方向接続を持っています。このようなつながりは、さらに複雑な形の感情学習につながり、行動に悪影響を与える可能性があります。たとえば、人は扁桃体を介して対の刺激を恐れるように条件付けされる可能性があります(たとえば、ショックと対になっているベルを怖がるようになる)。また、悪いことが起こった場所(教室でストレスを感じている場合など)を恐れるようになることもあります。 「コンテキスト」が条件付けに追加されると、海馬が学習に役割を果たす可能性があります。これは「状況による恐怖条件付け」と呼ばれ、条件付けされた場所に再び入ると、過剰で不適切な反応が生じる可能性があります。たとえば、教室での悪い経験をすべての教室に一般化することです。感情処理における扁桃体と海馬の相互作用の重要性は、気分障害や不安障害の素因となるリスクとして、扁桃体と海馬が不安気質に寄与していることを示す最近の研究で強調されている。霊長類では、不安気質に寄与する海馬の代謝活動は遺伝するが、扁桃体の活動は遺伝しない。これは、この行動表現型において、さまざまな脳領域に影響を与える遺伝子と環境のさまざまな役割を示唆しています。
覚えておくべき重要な点は、恐怖によって生み出された記憶にはさまざまな要素があり、これらのさまざまな要素が記憶をどのように制御または消去できるかに影響を与えるということです。興味深いことに、齧歯動物を対象とした研究では、動物は安全信号や悪いことが起こらない環境を認識するように条件付けされることもあることを示しています(例えば、決してショックが起こらない場所に条件付けされる)。このタイプの学習は、人間における特定の形態の心理療法の効果に寄与する可能性があり、消去学習や条件付き抑制だけでなく脱感作(脱条件)療法の基礎を形成するのに役立つ可能性があります。重要なのは、そのような学習は必ずしも「学習していない」というわけではなく、扁桃体、線条体、そしておそらく海馬が関与する別のタイプの学習である可能性が高いということです。
扁桃体は PFC とも接続しており、腹側 (眼窩) PFC および内側 (正中線) PFC と強いつながりがあります。扁桃体が背外側PFCの作業記憶に関与する領域と強いつながりを持たないことに注目するのは興味深いことであり、意識的な思考が扁桃体の機能にとって実際には重要ではないという考えを再び強調しています。実際、扁桃体が制御されているとき、作業記憶は一時的に停止されます。不安レベルが高まっているときに試験で良い成績を収めるのがどれほど難しいかを考えてみましょう。私たちが普段よく知っていることでも、不安がコントロールされているときには意識的に思い出せないことがあります。しかし、側方PFCと作業記憶は扁桃体を無効にすることができます(つまり、恐怖から抜け出す方法を考えることができます)。作業記憶と扁桃体の処理との関係、扁桃体とその接続が学習装置として機能する能力は、精神医学に大きな意味を持ち、気分障害や不安障害の治療に使用される心理療法の基礎となっている。
他の感情と他の脳領域
扁桃体は、感情処理において指揮者の役割を果たすと考えられている分散皮質下の脳システムである「大脳辺縁系」の一部にすぎません。大脳辺縁系の概念は時々議論されますが、それは皮質下システムが感情に関与していないからではなく、何が「大脳辺縁」構造を構成するかを定義することが複雑な接続性に基づいて困難になる可能性があるためです。扁桃体は感情処理において重要な役割を果たしますが、関係する唯一の構造ではありません。また、扁桃体が恐怖以上の処理に関与していることも明らかです。たとえば、扁桃体は怒り(「戦うか逃げるか」反応の「戦い」の部分)において重要な役割を果たしており、性的行動を含むポジティブな感情を引き起こすこともあります。扁桃体の異なる核が異なる一次感情に関与している可能性があります。内側核と後核は性的指向の感情や行動の処理に関与していると考えられていますが、前述したように外側核と中央核は恐怖の処理に関与しています。扁桃体、特にその内側部分が、攻撃的な行動を制御していると思われる基本的な「脅威」システムの一部であるという証拠もあります。
重要なことは、扁桃体自体は、終末線条床核(BNST)、線条体(特に腹側線条体)、下垂体を介したホルモン出力を調節する視床下部核、および脳幹を含む拡張システムの構成要素の 1 つにすぎないということです。モノアミン神経伝達物質の出力を調節する核。これらの脳幹核の例には、青斑核 (ノルエピネフリン合成部位)、背側縫線核 (セロトニン)、および腹側被蓋野 (ドーパミン) が含まれます。分散型感情処理ネットワークにおけるこれらの構造の接続と関与は、カテコールアミン、インドールアミン、ペプチド伝達物質、ホルモンが精神疾患にどのように関与しているかを理解するための枠組みを提供します。興味深いことに、げっ歯類と人間を対象とした研究では、恐怖よりもトリガーが明確ではない、より持続的な不安状態である不安は、BNSTを含む広範な扁桃体ネットワークに関与する原始的な急性恐怖反応とは異なることが示唆されています(図4を参照)。 4)。また、側坐核とドーパミンが感情処理に関与していることは、感情が動機にどのように関係しているかを理解するためのリンクを提供します。これについては後ほど詳しく説明します。
恐怖処理ネットワークに関する研究は、十分に特徴付けられた恐怖条件付けのげっ歯類モデルが存在するという事実によって大いに助けられてきました。これにより、解剖学的経路とシナプス機構をかなり詳細にマッピングできるようになりました。他の感情に対する同様のモデルはあまり開発されていません。それにもかかわらず、皮質の特定の領域が感情の処理と感情の意識的な認識の生成に関与していることを示す証拠が増えています。これらの皮質領域には、前島皮質 (AIC)、吻側前帯状皮質 (ACC)、および体性感覚皮質が含まれます。 AIC と ACC は、感情システムからマルチモーダルな入力を受け取り、感情システムと連携して機能する、より深い皮質構造です。これらは、嫌悪感、痛みの感情的要素、モチベーションなど、いくつかの感情を意識的に認識するために重要であると考えられます。嫌悪感は特に興味深い例です。この一次感情は、知覚に対する内臓(腸レベル)の反応を反映しており、環境内の何かが食べられるか有害かを評価するメカニズムとして進化した可能性があります。この「直感」は、最終的には「善悪」(私たちの道徳基準)を決定するなど、社会的に重要な別の意味を持つようになった可能性があります。言い換えれば、私たちが嫌悪感を感じるレベルは、それを超えると行為を続行できないレベルである可能性があります。道徳的意思決定に関する人間の神経画像研究は、これらの決定における私たちの最初の評価には、嫌悪感に関係する脳領域を含む感情ネットワークが関与していることを強く示唆しています。 AIC はこれらの経験において重要な役割を果たしており、恒常性状態 (自律神経系の状態、ホルモンレベル、覚醒状態)、感情状態、快楽状態、および社会的状況に関する情報をまとめることに熟達しています。道徳基準が感情の処理と密接に関連しているという概念は、認知システムと感情システムの驚くべき統合を示しています。図 4-5 は、人間の脳内の AIC の位置を示しています。
感情処理に関するこの議論の締めくくりとして、視床の下に位置する間脳の一部である視床下部の重要な役割にも焦点を当てたいと思います(これが名前の由来です。場所については付録を参照してください)。この領域は、扁桃体と同様、複数の小さな核の集合体であり、事実上、下垂体を介して脳と内分泌系を接続しています。さまざまな視床下部核は、食物摂取、代謝、性行動、概日リズム、ストレス反応、自律神経系(血圧、呼吸、体温)など、多くの行動プロセスおよび恒常性プロセスを調節しています。母性や社会的帰属などのさらに複雑な行動は、視床下部によって調節されるホルモン(オキシトシンなど)と強い関係があるようです。視床下部の機能とその制御の詳細は複雑で、15 以上の亜核が関与しています。視床下部の重要性は、一部の神経科学者が視床下部の働きを、要求に応じて身体機能を調整するサーモスタットの働きに例えているという考えによって強調されています。他の科学者はさらに一歩進んで、高次脳の大部分が視床下部を制御下に置くように設計されていると示唆しています。行動や精神障害に対する視床下部の重要性は、どれだけ強調してもしすぎることはありません。近年、いくつかの病気は視床下部の機能不全に直接関係していると考えられています。明確な例は、睡眠発作と脱力発作(筋緊張の喪失)を特徴とする疾患であるナルコレプシーが、ペプチドオレキシン(ヒポクレチンとしても知られる)を放出する特定の視床下部ニューロンの喪失に関与しているという発見である。今後、他の精神神経疾患でも同様の所見が発見される可能性がある。
————————図 4-5
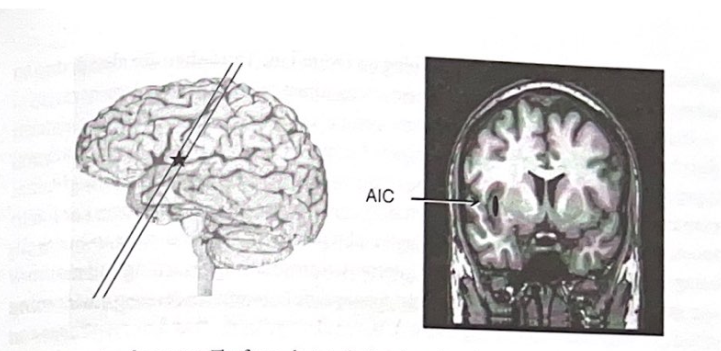
AIC
図 4-5 島皮質。図は島皮質の位置を示しています。この領域は、いくつかの感情のより高いレベルの処理において役割を果たします。略称:AIC(前島皮質)。 (許可を得て、Damasio、2005 年から翻案。)
————————-
何が脳の感情反応を引き起こすのでしょうか?
感情は私たちの精神生活の非常にダイナミックで強力な側面です。彼らは私たちが生き残るために制御を握ることができます。したがって、これらの反応を引き起こす刺激は、私たちが素早く認識するために重要であると思われます。人間の感情を引き起こすものについてはまだ学ばなければならないことがたくさんありますが、効果的なトリガーには、私たちの脳の「エラー」検出、つまり刺激が適切かどうかの迅速な(そして必ずしも意識的または正確ではない)評価が含まれているようであるという証拠が増えています。またはアクション)期待に応えられるかどうか。期待と知覚(または経験)結果の間に不一致がある場合、感情システムが活性化されて迅速な調整が行われることがあります。ご想像のとおり、最も激しい感情は否定的な評価によって引き起こされます。さらに、感情を活性化する計算は非常に粗雑である可能性があり、おそらく知識以上のものを反映していない可能性があります。
精神医学と臨床神経科学は、私たちの周りで何が起こっているかを推測(予測)します。したがって、これらはエラーを起こしやすく、不適切な応答を調整するために独自のエラー修正が必要になります。
認知神経科学者であり精神科医であるグレゴリー・バーンズは、著書『Iconoclast』の中で、人間の不安を引き起こすもののカテゴリーについて論じています。バーンズは人間のいくつかのタイプの恐怖について説明しており、彼の評価は、他の否定的な感情 (怒り、悲しみなど) がどのように活性化されるかにも関連する可能性がある興味深い思考の材料を提供します。最も基本的なレベルでは、恐怖は認識された害、つまり私たちの幸福に対する脅威によって引き起こされます。これらはかなり単純な解釈ですが、学習と記憶により、これらの反応は、あまり適切ではない状況(たとえば、実際には私たちに害を及ぼす可能性のないものに対して恐怖を抱くようになるなど)に一般化される可能性があります。バーンズはまた、恐怖反応は喪失や失敗の認識によって引き起こされる可能性があり、多くの場合、個人的な屈辱に対する恐怖を伴う可能性があると指摘しています。これは、拒絶されることや、ソーシャルネットワークから孤立して排除されることへの恐怖につながる可能性があります。人間は高度に社会的な動物であり、「群れ」との接触を失うことは私たちの幸福に悪影響を及ぼします。バーンズが説明した恐怖反応の最後のトリガーは、最も興味深いものであり、私たちの脳が抽象的な情報をどのように処理するかについて最も深い洞察を提供するものかもしれません。それは、未知のものに対する恐怖です。私たちの脳は不確実性にあまり耐えられないという概念をすでに強調しました。これは特に私たちの左半球に当てはまり、物事が一貫していないように見える場合、左半球は答えを作り上げる傾向があります。同様に、曖昧さと、ほとんどの決定には明確な正解か不正解があるという事実に対処することは、PFC にとって大きな課題です。確実性がない場合には、感情システムを使用して、より決定的な応答を提供することができますが、必ずしも正しい応答や有用な応答であるとは限りません。一部の認知科学者は、曖昧さへの対処の困難は、高次脳中枢に関わる深刻な機能障害の初期の症状の 1 つである可能性があると考えています。繰り返しになりますが、これは必ずしもこれらのセンターに病理があることを意味するものではなく、直面しているデータに苦戦しているということだけを意味します。
神経科学の観点から見ると、なぜ異なる状況下で異なる感情が引き起こされるのか、あるいはなぜ同じ人でも比較的似た状況に直面したときに異なる感情が引き起こされるのかは明らかではありません。また、同じような状況に直面したときに、なぜ不安を感じやすい人もいれば、怒りや悲しみを感じやすい人もいるのかもわかっていません。あるレベルの分析では、知覚された(または現実の)結果が期待を満たさないという計算は、その知覚が害をもたらすものである場合には不安を引き起こし、その知覚が損失または敗北を表すものである場合には悲しみを引き起こし、あるいは、被害をもたらすものである場合には怒りを引き起こす可能性がある。認識は不公平の一つです。個人をこれらの決定のいずれかに偏らせる要因は、私たちの動機付けシステムがどのように機能するか、そして私たちの期待が私たちの脳によってどのように決定されるかに大きく関係している可能性があります。
それは私たちの報酬の概念と私たちの期待を決定づける要因と結びついています。動機付け (および報酬) システムは、感情システムと同様、進化の観点から見ると古く、皮質下の感情システムと密接に絡み合っています。たとえば、Jaak Panksepp のスキームでは、「探索」と呼ばれる感情は、動物を環境探索に駆り立てる計算を伴うため、動機と考えられるものの一部を反映している可能性があります。また、特定のインセンティブが私たちの興味をそそる方法には、感情の活性化が含まれる場合があります。これらのインセンティブは生得的なものと学習によるものがあります。基本的な衝動(食べ物、性別、生存など)のように見えるものもあれば、より複雑で学習や環境に由来するものもあります。興味深いことに、人間には、自分自身の抽象的思考をインセンティブとして (または少なくとも動機付けとして) 使用する独特の能力があります。リード・モンタギューが指摘したように、抽象的な信念、たとえば宗教や政治的思想のために死をも厭わない動物はおそらく人間だけでしょう。彼はこれを「サメはハンガーストライキをしない」とうまくまとめています。後で詳しく説明しますが、ほとんどすべての乱用薬物は、神経伝達物質ドーパミンへの影響を介して、私たちの動機付けシステムの活動を調節し、奪います。
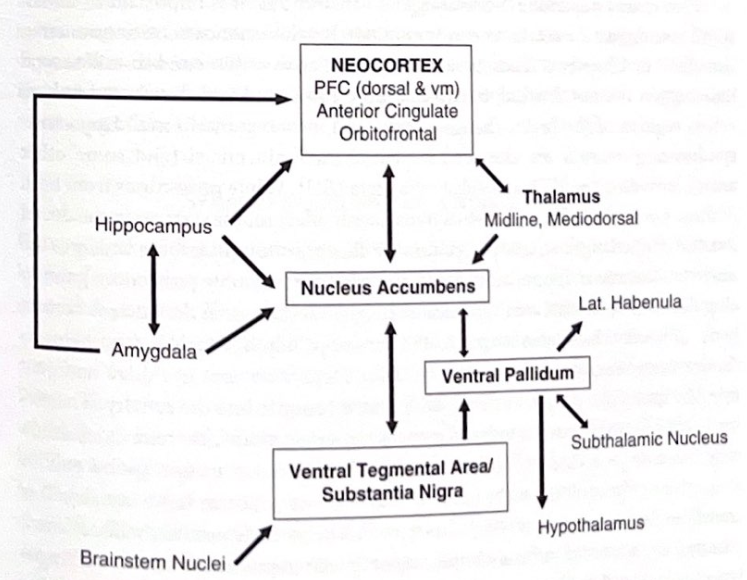
私たちの動機付けシステムには、側坐核、腹側淡蒼球、中脳腹側被蓋野 (VTA) を含む皮質下ネットワークが関与しています (図 4-6)。このシステムは、PFC を含む視床皮質システムと強く相互作用し、動機と報酬の両方を計算できるように見えますが、これらは処理の観点からは必ずしも同じものではありません。 VTA はドーパミンを合成する領域であり、VTA からの突起によって放出されるドーパミンは側坐核の重要な調節因子です。ドーパミンの影響は、側坐核を超えて、エラー検出に重要な吻側帯状皮質の接続領域や、作業記憶を調節するPFCの領域にも広がります。画像研究に基づくと、報酬の予期は側坐核とVTAを活性化するようですが、報酬を経験すると内側尾状核、被殻、そして最終的には背側尾状核と補足運動野も関与するようです。ドーパミンは、初期反応を促進し、行動の結果に関与するより分散された回路を動員するのに役立ちます。
————————図 4-6
海馬
扁桃体
NEOCORTEX PFC (背側および vm) 前帯状皮質 眼窩前頭
視床正中線、中背筋
側坐核
腹部淡蒼球
何年も。手綱核
モチベーション: 目標を持つことの重要性
モチベーションはどのように機能するのでしょうか?
モチベーションはルドゥーのメンタル三部作の 3 番目の要素です。これには、目標を設定して達成する方法を決定する計算が含まれます。モチベーションは身近にある
精神医学と臨床神経科学
脳幹核
視床下核
腹側被蓋野/黒質
視床下部
図 4-6 報酬回路。この図は、腹側被蓋野、側坐核、新皮質の領域の中心的な役割を強調して、動機付けシステム内の主要な構造と接続を示しています。キー構造の場所については、付録を参照してください。
————————-
モチベーションシステムはどのように機能するのでしょうか?側坐核は、目標、感情、記憶に関する入力を統合し、腹側淡蒼球を介して作用する信号を生成して、モーターの出力と行動を促進する重要な領域であると考えられます。このシステムでは、ドーパミンがエラー検出信号として重要な役割を果たしているようです。ドーパミンは報酬信号であると考える人もいますが、現在の考えではドーパミンが異なる役割を果たすことが示唆されています。ドーパミンは、認識が期待どおりかどうかを判断するのに役立ち、ネットワーク内の他の領域が重要な(顕著な)活動に注意を向けるのに役立ちます。これが実際に意味することは、VTA におけるドーパミン ニューロンの発火は、観察と期待の計算による比較を反映しているということです。結果が予想より良好な場合、VTA のドーパミン ニューロンは活動電位の発火率を高め、その結果、側坐核、帯状皮質、および PFC でより多くのドーパミンが放出されます。結果が期待を満たさないと、興奮が減り、ドーパミンレベルが低下します。したがって、モンタギューの言葉を借りれば、これらの特定のニューロンの発火とドーパミンの放出が「批判信号」として機能します。
ドーパミンはなぜ「やる気を起こさせる」のでしょうか?これに答えるには、ドーパミンが神経伝達物質としてどのように作用するかを理解することが重要です。第 3 章で説明したように、これは遅いモノアミン送信機です。したがって、その動作によってミリ秒単位の情報転送が行われるわけではありません。代わりに、脳の領域全体にわたってより長期的かつ分散的に作用し、領域間および領域内の「調子」を変化させます。ドーパミンを合成するニューロンは、VTA や黒質 (SN) を含むいくつかの中脳核 (およびその他の領域) に集中しています。これらの核の両方からの投射は拡散していますが、SN 投射は主に背側線条体の運動系に影響を与えます (黒質線条体投射と呼ばれます)。 VTA 投影は、側坐核、海馬、扁桃体 (中脳辺縁系投影)、および帯状皮質および前頭前皮質 (中皮質投影) に広く分布しています。側坐核では、ドーパミンが腹側淡蒼球への出力を促進し、それが運動活動や行動の変化を促進します。ドーパミンは(グルタミン酸のように)それ自体でニューロンの発火を引き起こすのではなく、むしろ、強い入力に応答して発火するように、受信(シナプス後)ニューロンの活動にバイアスをかけているようです。したがって、ドーパミンは、ニューロンに強い入力にのみ反応するように指示する一種の「音量制御」として機能する可能性があります(つまり、ニューロンはそれらの信号に注意を払う必要があります)。これは、ドーパミンが「顕著性」(つまり、入力が注目に値するほど重要であるかどうか)を決定するのに役立つ要素として機能することに変換されます。吻側帯状皮質では、このシグナル伝達はエラー予測を媒介する一方、外側PFCでは注意と作業記憶を集中させるのに役立ちます。 PFC では、これは一種の「フィルタリング」機能とみなすこともできます。ドーパミンが多すぎる、または少なすぎる病的状態では、脳は刺激が重要であるか無関係であるかを判断するのが困難になることがあります。これは統合失調症患者にとって大きな問題となる可能性があり、ドーパミン伝達の変化が作業記憶や認知の欠陥に寄与すると考えられている。
動機付けシステムにおけるドーパミン伝達は、目標の更新と選択に役立つ経験依存の処理の一種である強化(インセンティブベース)学習にも重要であるようです。テリー・セジノウスキー氏らによると、強化学習は、古代の予測システム(ドーパミン)を使って現代世界と関わり、その世界から何がうまくいき、何がうまくいかないのかを学ぶことを可能にするメカニズムだという。モンタギューは、このプロセスを一連の計算ステップとして説明します。まず、(内部または外部の)知覚により、長期記憶に保存された値関数と比較される即時信号が生成されます。この比較により、前述したように「臨界信号」(ドーパミン細胞の発火)が発生します。次に、批判信号の積 (ドーパミン) を使用して選択を導き、受信ニューロンの反応にバイアスをかけることで次の目標を選択します。このいわゆる「報酬予測誤差信号」は、その後の意思決定にバイアスをかけるだけでなく、経験から何を学び、将来の期待を設定するために使用されるかにも影響を与えます。この経験の学習部分は海馬の関与を反映している可能性が高く、海馬ではドーパミンの入力が調節の役割を果たし、特定の長期記憶の生成に重要であると考えられます。人間を対象とした最近の画像研究は、脳がエラー検出と報酬処理をどのように区別するかを理解することに焦点を当てています。エラーは、認識されたエラーが私たちによってなされたものであるか他人によってなされたものであるかに関係なく、後内側 PFC (前補足運動野および吻側帯状帯を含む) を強く活性化するようです。後者は、私たちが他人の行動からどのように学ぶかを示す一例です。対照的に、線条体 (特に腹側線条体) の活動は、エラー状態に関係なく報酬によって行われます。
私たちの期待は何によって決まるのでしょうか?
期待と結果の比較は強化学習において重要な役割を果たしますが、何が私たちの期待を設定するのでしょうか?明らかに、それらは静的ではなく、多くの場合コンテキストに依存します。したがって、それらは私たちの感情や記憶からの修正を受ける可能性があります。それにもかかわらず、私たちの期待は、多くの場合、私たち自身の中核的で長年培ってきた側面、つまり「性格」という広い見出しに分類される側面を反映しています。性格とは、私たちが自分自身や世界についてどのように考えるかについての永続的なパターンを指します。それはさまざまな方法で説明できますが、脳システムの観点から役立つと思われる方法の 1 つは、C. ロバート クローニンジャーによって支持されています。クロニンジャー氏によれば、人格には気質と性格という 2 つの主要な側面があります。気質は、私たちが世界とどのように関わり、世界に反応するかに影響を与える基本的な習慣とスキルを反映しています。クローニンジャーは、気質の 4 つの側面、すなわち新奇性の追求、危害の回避、報酬への依存、および持続性を定義しています。性格特性は、私たちが世界にどのように参加するかを決定します。これには、協調性、自主性、自己超越性が含まれます。
遺伝は人の気質と性格特性の決定に大きな役割を果たしますが、両方とも経験によっても変化します。気質特性は、私たちの注意を引くものや私たちにとって重要なものを決定するのに特に重要であるようです。したがって、これらの特性は顕著性を判断し、私たちの内部の期待のバロメーターを設定するのにも役立つようです。私たちは物事が静的なときと変化しているときのどちらが最も幸せですか?私たちはリスクの高い仕事よりも安全を優先しますか?他人の承認は私たちにとってどれくらい意味があるのでしょうか?神経画像研究は、これらの特性に寄与し、促進する神経系の解明に役立っています。たとえば、新奇性の追求には海馬、扁桃体、側坐核を含むネットワークが関与しているようですが、報酬依存には PFC や線条体の領域が関与しているようです。持続には、側坐核だけでなく、外側眼窩皮質および内側前頭前皮質も含まれます。これらの形質における側坐核の役割は、動機付け、誤り検出、強化学習における役割を考慮すると興味深いものである。また、新奇探求の研究では、探索的傾向が高い人は、腹側中脳ドーパミンニューロンにおける特定のクラスのドーパミン受容体(D2受容体)の発現が低下していることが示されています。これらの個人は、新規性の追求が低い個人と比較して、新規の刺激に対するドーパミン反応が強化されています。
これらの観察は、精神障害におけるパーソナリティの役割や、パーソナリティ障害を持つ人が自分の気質や性格から生じる困難に対処するための潜在的な戦略を考える上で重要です。クローニンジャーによれば、性格特性の欠陥はパーソナリティ障害の可能性を示す特徴であり、気質特性の違いがパーソナリティ障害の形態を決定するという。たとえば、非協力的な行動や自主性の欠如は、多くのパーソナリティ障害で観察されます。しかし、これらの性格上の問題に直面した場合の新規性の追求が高く、危害回避性が低いことは、反社会的行動(嘘をつく、学校や仕事の成績が悪い、性的乱交、喧嘩など)を示すことと相関している。同様の分析は他のパーソナリティ障害にも適用できます。
私たちは、モチベーションが脳の機能において果たす重要な役割と、モチベーション、認知、感情がどのように密接に絡み合っているかをさらに強調して、この議論を終えたいと思います。カール・フリストンは、私たちは皆、世界の内部表現を持っており、それと比較して、期待に沿うかどうかを判断するという仮説を立てました。この保存された「世界モデル」に基づくモチベーションは、脳が障害(および熱力学の第二法則)を迅速に克服するのに役立つと彼は主張しています。これは、入力間の偏った注意と競争の基礎を形成し、上昇する変調器 (感情) によって割り当てられた値によって、入力の処理方法と出力の生成方法を制御できるようになります。これは、実行機能を研ぎ澄まし、脳への「驚き」(高次認知機能にとっての悩み)を最小限に抑えるのに大いに役立ちますが、サンプリングされる内容とサンプリング方法に使用される戦略と同様に、サンプリングされる内容とサンプリング方法に偏りがあることも保証します。処理。後者の例としては、報酬に大きく依存している人は、パフォーマンスに対して報酬が与えられる環境下では作業記憶のパフォーマンスがより大きく向上するという発見がありますが、興味深いことに、作業記憶の向上は通常、直接報酬が得られないタスク中に発生します。 。彼らは、報酬の可能性を高める積極的な認知制御戦略 (この章の前のセクションを参照) を採用することでパフォーマンスを向上させます。これは、モチベーションと認知がどのように絡み合っているかを改めて浮き彫りにします。
モチベーションの食欲(アプローチ)の側面は主に腹側線条体回路によって駆動されます。モチベーションには、主に扁桃体に基づく回路と、PFC ネットワークによって媒介される調節要素によって駆動される回避の側面があると考えることもでき、心の 3 つの要素がどのように絡み合っているかが改めて強調されています。
要約: 脳システムと心の簡単な概要
私たちは、相互に関連する脳ネットワークからなる三部作として心を概念化することは、精神科の症状や障害について考える上で発見的に価値があると信じています。これらの点を単純化しすぎる危険を承知で、この章で説明した内容のより基本的な要約を提供してこの議論を終了します。確かに、以下の説明は科学的および脳処理の観点からすると洗練されていませんが、このような単純化は臨床医が脳システムが精神医学的問題にどのように寄与しているか、またそれらの問題に治療的にどのようにアプローチするかを実際に考えるのに役立つと考えています。
思考、感情、モチベーションの根底にある脳ネットワークは強く相互作用しており、これらの領域にわたる欠陥はすべての主要な精神疾患で発生します。意識的な思考は新皮質の権限であり、PFC はこのプロセスの中心です。それは意識的な脳で起こるすべてのことの司会者であり、意思決定者であり、解釈者です。 PFC はそれ自体ですべての問題を解決できるわけではありませんが、脳内のどこの問題を解決できるかを最適に制御します。他のスモールワールド ネットワークが自己組織化して動作する方法と同様に、PFC には非常に高度に接続されたノードが含まれており、他の多くの脳領域に影響を与える長距離接続を備えています。また、他のシステムから欠陥のある情報を取得すると、機能不全やエラーが発生する可能性があります。
扁桃体は感情の処理に重要です。これは、課題を検出し、視床下部やその他の恒常性システムと連携して個人に警告し、適切と思われる場合には迅速に問題を修正する「番犬」のようなものと考えられるかもしれません。ドーパミン-側坐核システムは、PFC の重要な部分とともに、モチベーションにおいて重要であり、行動の「サーモスタット」に似ていると考えることができます。期待と結果の間に不一致が発生した場合、このシステムは脳がコンパスを調整してリセットするのに役立ちます。海馬については後の章で扱いますが、今のところは、収束入力の処理における海馬の重要な役割を理解することが重要です。それは「統合器」のように機能し、私たちの記憶や感情を含む多くのものを結び付け、その結果、経験に基づいたより豊かな連想や記憶が生まれます。処理されたほぼすべての情報は最終的に海馬に到達し、海馬は独自の処理を複数の脳システムと共有できるようになります。海馬は新しさの検出にも重要であるため、「新しい」と考えられるものは最終的にこのシステムに入ります。最後に、前島皮質と吻側帯状皮質は、内受容(内部)表現と主観的経験を意識にもたらし、行動に影響を与えるのを助ける「通訳者」として重要な役割を果たします。これら 2 つの領域は、より最近に進化した PFC に対する古代の進化システムの活動を解釈するのに役立ちます。いくつかの重要な領域を取り上げましたが、一般的な ICN の概念を念頭に置くことも重要です。これらの領域は孤立して機能するのではなく、すべて相互作用して精神処理を導く分散型脳回路の一部です。さらに、脳システム内および脳システム間の相互作用の複雑さについての私たちの理解はまだ初期段階にあり、新しい情報が得られるにつれてシステムの神経科学が進歩しています。たとえば、小脳には全脳ニューロンの 50% 以上が含まれており、主に運動機能を調節していると考えられてきました。いくつかの証拠は、これは単純すぎることを示唆しており、この領域は言語、気分、および実行制御の基礎となるネットワークにも参加している可能性があります。
これらの脳システムはコンピューターやカメラに似たものを生み出すものではないことを理解することも重要です。彼らはエネルギー節約のための近道を選択することで間違いを犯しますが、健康な脳では、それらの間違いを修正する素晴らしい仕事をし、一貫した思考、感情、行動につながります。これは、私たちの脳が古代のシステム (扁桃体と側坐核) に中間のシステム (海馬) と比較的現代的なシステム (新皮質) を組み合わせているという事実を考慮すると、さらに驚くべきことです。デヴィッド・リンデンは著書『The Accidental Mind』の中で、そのような取り決めを「クラッジ」と呼んでいます。この用語は、大まかに言うと、何らかの形で仕事を成し遂げるように見える、断片的にまとめられたものの束を意味します。工学的な観点から見ると、これが優れた計算デバイスを設計する最適な方法であるかどうかは明らかではありませんが、このシステムは、パターン認識、抽象化、概念生成の点で、最も洗練されたコンピューターよりも優れた機能を備えています。それがうまく機能するのは驚くべきことです…ほとんどの場合。
————————-
留意事項
精神障害は脳の障害であり、心の認知、感情、モチベーションのあらゆる側面にわたる機能不全を反映しています。
特定の病気では、特定の症状が優勢である場合がありますが(気分障害や不安障害などの感情的な問題)、認知、感情、動機の問題は、ほとんどの場合併発しています。脳がグローバル ネットワークとして配線されているため、問題が発生した場合には、これら 3 つの領域にまたがる症状がほぼ確実に併発します。
脳は複雑な器官ですが、精神処理は効果的に説明できます
システム神経科学とICNの観点から理解されています。後続の章でさらに詳しく説明しますが、特定の ICN の欠陥は精神症状を予測することができ、リハビリテーションの取り組みや心理療法の対象となる可能性があります。分子機構の欠陥は、特定の薬理学的介入の影響を受けやすい可能性が高くなります。
推奨される読み物
Buckner, R.L.、Andrews-Hanna, J.R.、Schacter, D.L. (2008)。脳のデフォルト ネットワーク: 解剖学、機能、病気との関連性。ニューヨーク科学アカデミー年報、1124 年、
1-38。
ルドゥー、J. (2002)。シナプス自己: 私たちの脳がどのようにして私たちになるのか。ニューヨーク:バイキング・プレス。モンタギュー、R. (2006)。なぜこの本を選んだのでしょうか?私たちがどのように意思決定を行うか。ニューヨーク:ダットン・プレス。
その他の参考文献
バーンズ、G. (2008)。 Iconoclast: 神経科学者が、これまでとは異なる考え方をする方法を明らかにします。ボストン:ハーバード大学ビジネスプレス。
J. W. ビスリー (2011)。視覚的注意の神経基盤。 Journal of Physiology (ロンドン)、589、49-57。
ブレア、R.J.R. (2010)。精神病質と反社会的行動の神経画像化: 的を絞ったレビュー。現在の精神医学レポート、12、76-82。
Bledowski、C.、Rahm、B.、および Rowe、J.B. (2009)。ワーキングメモリでは何が「機能」するのでしょうか?重要な情報の選択と更新のための個別のシステム。神経科学ジャーナル、29、735-741。
Braver, T.S.、Paxton, J.L.、Locke, H.S.、および Barch, D.M. (2009)。人間の前頭前野における認知制御の柔軟な神経機構。米国科学アカデミー紀要(米国)、106、7351-7356。
Casey, B.J.、Duhoux, S.、および Cohen, M.M. (2010)。思春期: 伝達、移行、翻訳は思春期とどのような関係があるのでしょうか?ニューロン、67、749-760。
Cloninger、C.R. (2004)。良い気分: 幸福の科学。ニューヨーク:オックスフォード大学出版局。
Cohen, M.X.、Schoene-Bake, J.-C.、Elger, C.E.、Weber, B. (2009)。人間の線条体の接続性に基づく分離により、性格特性が予測されます。ネイチャー・ニューロサイエンス、12、32-34。
A.R.A.コンウェイ、M.J.ケイン、R.W.エングル(2003)。作業記憶容量とその一般的な知能との関係。認知科学の動向、7、547-552。
Corbetta, M.、Patel, G.、Shulman, G.L. (2008)。人間の脳の方向転換システム: 環境から心の理論まで。ニューロン、58、306–324。
ダマシオ、A. (1999)。何が起こったのかという感覚:意識の形成における身体と感情。カリフォルニア州サンディエゴ:ハーコート。
Davis, M.、Walker, D.L.、Miles, L.、Grillon, C. (2010)。ラットとヒトにおける一時的恐怖と持続的恐怖: 恐怖と不安における拡張扁桃体の役割。神経精神薬理学レビュー、35、105-135。
de Bruijn, E.R.A.、de Lange, F.P.、von Cramon, D.Y.、Ullsperger, M. (2009)。エラーにやりがいがあるとき。神経科学ジャーナル、29、183-186。
Dosenbach, N.U.F.、Fair, D.A.、Cohen, A.L.、Schlaggar, B.L.、Petersen, S.E. (2008)。トップダウン制御のデュアルネットワークアーキテクチャ。認知科学の動向、12、99-105。フリストン、K. (2010)。フリーエネルギー原理: 統合された脳機能?自然のレビュー
神経科学、11、127-138。
ガザニガ、MS (2008)。人間: 私たちをユニークにする背後にある科学。ニューヨーク:ハーパーコリンズ。
ゴールドバーグ、E. (2001)。経営者の脳: 前頭葉と文明的な精神。ニューヨーク:オックスフォード大学出版局。
グレイ、K.、ジェンキンス、A.C.、ヘバーライン、A.S.、ウェグナー、D.M. (2011年)。精神病理学における心の認識の歪み。米国科学アカデミー紀要(米国)、108、477-479。
Greene, J.D.、Nystrom, L.E.、Engell, A.D.、Darley, J.M. & Cohen, J.D. (2004)。道徳的判断における認知的葛藤と制御の神経基盤。ニューロン、44、389-400。 Gusnard, D.A.、Ollinger, J.M.、Shulman, G.L.、Cloninger C.R.、Price、J.L. Van Essen, D.C.、他。 (2003年)。持続性と脳の回路。米国科学アカデミー紀要(米国)、100、3479-3484。
Haber, S.N.、Knutson, B. (2010)。報酬回路: 霊長類の解剖学と人間のイメージングのリンク。神経精神薬理学レビュー、35、4-26。
J. ホーキンス (S. ブレークスリーとともに)。 (2004)。知性について。ニューヨーク:タイムズブックス。
彼、J.J.、シュルマン、G.L.、スナイダー、A.Z.、コルベッタ、M. (2007)。神経障害における神経伝達障害の役割。 Current Opinion in Neurology、20、655-660。 Javanovic, T.、および Ressler, K.J. (2010)。恐怖抑制の神経回路と遺伝学が、PTSD についての私たちの理解をどのように伝えるのか。アメリカ精神医学ジャーナル、167、648-662。 Jimura, K.、Locke, H.S.、および Braver, T.S. (2010)。報酬を与える動機付けの文脈における認知強化の前頭前皮質の仲介。米国科学アカデミー紀要(米国)、107、8871-8876。
Khalsa, S.S.、Rudrauf, D.、Feinstein, J.S.、Tranel, D. (2009)。内受容意識の経路。 Nature Neuroscience、12、1494-1496。
Kouneiher F.、Charron, S.、および Koechlin, E. (2009)。人間の前頭前野におけるモチベーションと認知制御。 Nature Neuroscience、12、939-945。
Laird, A.R.、Eickhoff, S.B.、Li, K.、Robin, D.A.、Glahn, D.C.、および Fox, P.T. (2009)。座標ベースのメタ分析モデリングを使用して、デフォルト モード ネットワークの機能的異質性を調査します。神経科学ジャーナル、29、496-505。
レイコフ、G.、ヌネス、R. (2000)。数学はどこから来るのか: 肉体化された心がどのようにして数学を実現するのか。ニューヨーク:基本的な本。
リンデン、D.E.J. (2007)。人間の脳のワーキングメモリネットワーク。神経科学者、13、257-267。
リンデン、D.J. (2007)。偶然の心: 脳の進化が私たちにどのように愛、記憶、夢、そして神を与えてきたのか。マサチューセッツ州ケンブリッジ:ベルナッププレス。
Liu, H.、Stufflebeam, S.M.、Sepulcre, J.、Hedden, T.、および Buckner, R.L. (2009)。人間の脳の非対称性が複数の要因によって制御されているという固有の活動からの証拠。米国科学アカデミー紀要(米国)、106、499-503。
Newberg, A.、D’Aquili, E.、Rause, V. (2001)。なぜ神は去らないのか。脳科学と信念の生物学。ニューヨーク:バランタイン・ブックス。
野村、E.M.、グラットン、C.、ヴィッサー、R.M.、カイザー、A.、ペレス、F.、およびデスポジート、M. (2010)。局所性脳損傷患者における 2 つの認知制御ネットワークの二重解離。米国科学アカデミー紀要(米国)、107、17-22。
Oler, J.A.、Fox, A.S.、Shelton, S.E.、Rogers, J.、Dyer, T.D.、Davidson, R.J. 他(2010年)。不安気質の扁桃体基質と海馬基質は、遺伝率が異なります。自然、466、864-868。
パンクセップ、J. (2004)。感情神経科学: 人間と動物の感情の基礎。ニューヨーク:オックスフォード大学出版局。
パンクセップ、J. (2006)。進化精神医学における感情エンドフェノタイプ。神経精神薬理学および生物学的精神医学の進歩、30、774-784。
Quartz、S.R.、Sejnowski、T.J. (2002)。嘘つき、恋人、ヒーロー: 新しい脳科学が明らかにする、私たちがどのようにして自分になるのか。ニューヨーク:ウィリアム・モロー。
レイクル、M.E.、スナイダー、AZ (2007)。脳機能のデフォルトモード: 進化するアイデアの簡単な歴史。ニューロイメージ、37、1083-1090。
Riley, K.P.、Snowden, D.A.、Desrosiers, M.F.、Markesbery, W.R. (2005)。幼少期の言語能力、晩年期の認知機能、および神経病理:修道女の研究からの発見。老化の神経生物学、26、341-347。
Shin、L.M.、および Liberzon, I. (2010)。恐怖、ストレス、不安障害の神経回路。神経精神薬理学レビュー、35、169–191。
Vann, S.D.、Aggleton, JP、Maguire, E.A. (2009)。脾後皮質は何をしているのでしょうか? Nature Reviews Neuroscience、10、792-802。
Zald, D.H.、Cowan, R.L.、Riccardi, P.、Baldwin, R.M.、Ansari, M.S.、Li, R.、他。 (2008年)。中脳のドーパミン受容体の利用可能性は、ヒトにおける新奇性を求める性質と逆相関しています。神経科学ジャーナル、28、14372-14378。
Ziemann、A.E.、Allen、J.E.、Dahdaleh、N.S.、Drebot、I.I.、Coryell、M.W.、Wunsch、A.M.、他。 (2009年)。扁桃体は、二酸化炭素とアシドーシスを検出して恐怖行動を誘発する化学センサーです。セル、139、1012-1021。精神医学と臨床神経科学
